Автором ассоциативной теории памяти является:
спросил от Akma в категории Тесты ЕНТ, КТА, ВОУД Ответы на тесты ЕНТВаш ответ
| Отображаемое имя (по желанию): |
| Отправить мне письмо на это адрес если мой ответ выбран или прокомментирован:Отправить мне письмо если мой ответ выбран или прокомментирован |
Конфиденциальность: Ваш электронный адрес будет использоваться только для отправки уведомлений. |
| Анти-спам проверка: |
Чтобы избежать проверки в будущем, пожалуйста войдите или зарегистрируйтесь. |

Похожие вопросы
Кто является автором «теории администрирования»?
спросил от Akma в категории Тесты ЕНТ, КТА, ВОУД Ответы на тесты ЕНТ
Автором метрической школы умственных способностей является:
спросил от Akma в категории Тесты ЕНТ, КТА, ВОУД Ответы на тесты ЕНТ
Кто является автором Успенского собора в кремле, выстроенном в 1479 г?
спросил от Akma в категории Тесты ЕНТ, КТА, ВОУД Ответы на тесты ЕНТ
Кто из скульпторов поздней классики в греческом искусстве является автором Галикарнасского мавзолея?
Автором какого первого гигантского архитектурного сооружения Египта является зодчий Имхотеп?
спросил от Akma в категории Тесты ЕНТ, КТА, ВОУД Ответы на тесты ЕНТ
Автором метода заучивания (метода последовательных воспроизведений) является:
спросил от Akma в категории Тесты ЕНТ, КТА, ВОУД Ответы на тесты ЕНТ
Кто является автором книги «5 законов библиотечной науки» ?
спросил от Akma в категории Тесты ЕНТ, КТА, ВОУД Ответы на тесты ЕНТ
Кто является автором естественнонаучной психофизиологической основы логопедии о взаимодействии первой и второй сигнальной системы?
спросил от Akma в категории Тесты ЕНТ, КТА, ВОУД Ответы на тесты ЕНТ
Автором романса «Соловей» является:
спросил от Айым в категории Общество
Автором записей, напечатанных под названием «Хождение за три моря», является
спросил от Zhanat в категории Города и Страны
Первые сведения о населении и хозяйстве Казахстана можно найти в «Сборнике летописей», автором которого является
спросил от Вопросы и ответы в категории Тесты ЕНТ, КТА, ВОУД Ответы на тесты ЕНТ
Мыржакып Дулатов является автором произведения
спросил от Вопросы и ответы в категории Школы
Мыржакып Дулатов является автором сборника
спросил от Вопросы и ответы в категории Школы
Методологической основой современной теории обучения является:
спросил от Akma в категории Тесты ЕНТ, КТА, ВОУД Ответы на тесты ЕНТ
Основным понятием в теории З. Фрейда является понятие:
Фрейда является понятие:
спросил от Akma в категории Тесты ЕНТ, КТА, ВОУД Ответы на тесты ЕНТ
- Все категории
- Авто-Мото 839
- Бизнес, Финансы 1,663
- Праздники 58
- Города и Страны 1,230
- Досуг, Развлечения 455
- Еда, Кулинария 231
- Животные, Растения 5,988
- Знакомства, Любовь, Отношения 71
- Искусство и Культура 10,186
- Игры 275
- Кино 41
- Музыка 513
- Компьютеры, Связь 2,305
- Красота и Здоровье 1,098
- Наука, Техника, Языки 3,265
- Ұстаз 1,073
- Образование 6,736
- Общество, Политика, СМИ 1,744
- Юридическая консультация 144
- Путешествия, Туризм 101
- Работа, Карьера 98
- Казахские традиции 25
- Семья, Дом, Дети 178
- Спорт 102
- Стиль, Мода, Звезды 33
- Товары и Услуги 4,279
- Фотография, Видеосъемка 357
- Логические задачи 266
- Тесты ЕНТ, КТА, ВОУД Ответы на тесты ЕНТ
28,735
- Тесты по географии с ответами 28
- Юмор 18
- Другое 14,198
7
Ассоциативная теория памяти — Учебник по общей психологии Юнита (Психология)
3. Ассоциативная теория памяти. Виды ассоциаций и законы их образования. Критика ассоцианизма
Ассоциативная теория памяти. Виды ассоциаций и законы их образования. Критика ассоцианизма
3.1. Понятие и виды ассоциаций
Учение об ассоциациях можно считать первой теорией памяти; его истоки прослеживаются уже в платоновских диалогах (“Федон”) и у Аристотеля (de mem. 2), а расцвет ассоцианизма приходится на XVIII в. Д. Юм, Д.Гартли), когда принцип ассоциации был распространен на всю область психического. В конце XIX – начале XX в. ассоциативная теория, объясняющая динамику процессов памяти принципом ассоциации, стала ведущим направлением в психологии. Основное понятие учения, ассоциацию, можно определить как связь между психическими явлениями, при которой актуализация одного из них влечет за собой появление другого. Предметы и явления запечатлеваются и воспроизводятся не изолированно, а в связи друг с другом, которая обусловлена их реальными объективными отношениями. Физиологической основой ассоциации служат нервные пути в коре мозга, с помощью которых протекают процессы запоминания и воспроизведения.
Со времен Аристотеля (считается, что именно он открыл законы образования ассоциаций) известны четыре вида ассоциаций. Ассоциации по смежности представляют собой установление связи между явлениями по признаку пространственно-временных отношений. В ассоциациях по сходству связь между явлениями устанавливается по признаку сходства, в ассоциациях по противоположности – по признаку контраста. Ассоциации по каузальности образуются по признаку причинно-следственных отношений. Ассоциации могут быть простыми, если они связывают между собой только два представления, и сложными, если одно представление объединяется со многими другими и образует ассоциативный комплекс (см. рис. 1, б).
3.2. Экспериментальные методы исследования памяти в ассоцианизме
В конце XIX в. немецкий психолог Герман Эббингауз, опираясь на ассоциативную теорию, разработал основные приемы изучения памяти с помощью объективных методов в противовес интроспективным методам школы В. Вундта. В своем классическом исследовании “О памяти” (1885) Г. Эббингауз дает следующее определение ассоциации: “Душевные образования называются ассоциированными, если они когда-либо раньше были пережиты вместе, и существует более или менее основательное допущение, что при существующих условиях они могут вызывать друг друга” ([3], с.10). Общее правило возникновения ассоциаций таково: “…Если какие-либо душевные образования когда-нибудь заполняли сознание одновременно или в близкой последовательности, то впоследствии повторение одних членов этого прежнего переживания вызовет представления и остальных членов, хотя бы первоначальные причины их и отсутствовали” ([3], с.9).
Вундта. В своем классическом исследовании “О памяти” (1885) Г. Эббингауз дает следующее определение ассоциации: “Душевные образования называются ассоциированными, если они когда-либо раньше были пережиты вместе, и существует более или менее основательное допущение, что при существующих условиях они могут вызывать друг друга” ([3], с.10). Общее правило возникновения ассоциаций таково: “…Если какие-либо душевные образования когда-нибудь заполняли сознание одновременно или в близкой последовательности, то впоследствии повторение одних членов этого прежнего переживания вызовет представления и остальных членов, хотя бы первоначальные причины их и отсутствовали” ([3], с.9).
Для экспериментального исследования памяти Г. Эббингауз предлагает использовать следующие методы:
I. Методы узнавания. Элементы материала, который заучивает испытуемый, располагаются в произвольном порядке среди новых, очень похожих на искомые. Испытуемый должен просмотреть их и идентифицировать те, которые он заучивает.
II. Методы воспроизведения. а) Метод запоминаемых членов – простейший в данной группе методов. Элементы запоминаемого материала предъявляются однократно и воспроизводятся непосредственно или спустя некоторое время после предъявления. Измеряется количество правильно названных элементов и время, затраченное на припоминание.
б) Метод заучивания. Ряд элементов предъявляется многократно и заучивается до достижения определенного критерия, например до первого безошибочного воспроизведения. Фиксируют число проб (повторений) и время, затраченное на заучивание. По результатам строится кривая научения: по оси абсцисс откладывается число проб, а по оси ординат – число элементов, правильно воспроизведенных в каждой пробе (подробнее о кривых научения см. раздел 5).
в) Метод антиципации. Элементы материала группируются в ряды аbcd.., и предъявляются один или несколько раз. Испытуемый пытается воспроизвести их, соблюдая установленный порядок. В случае ошибки или пропуска экспериментатор называет искомый элемент.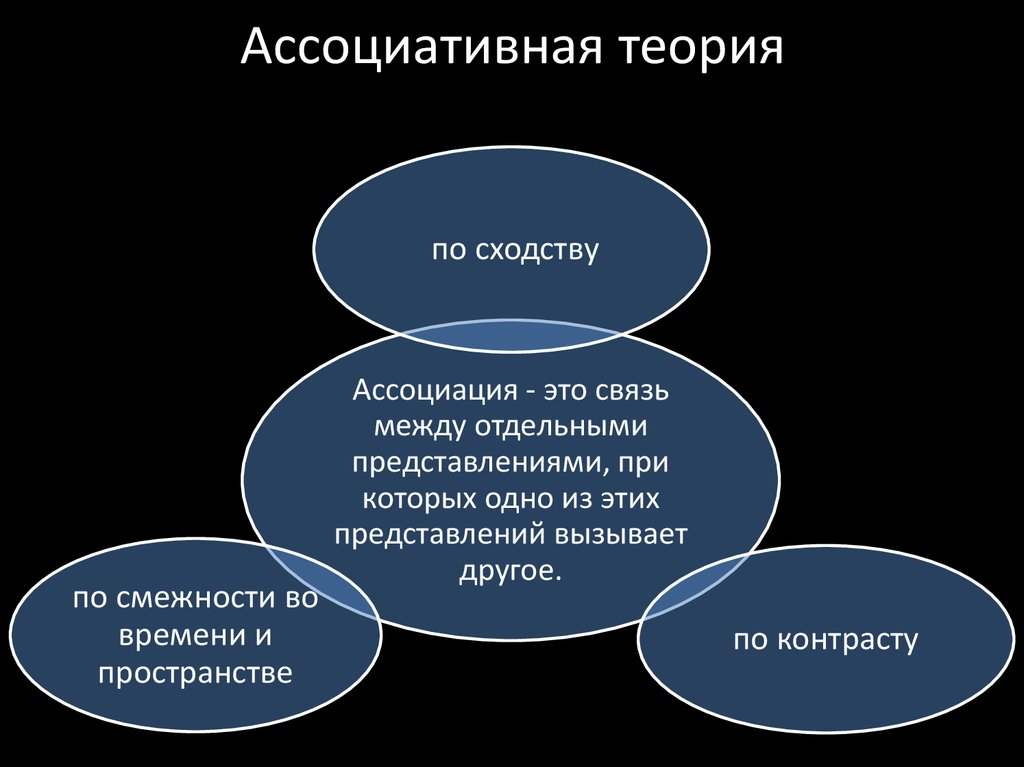 Процедура продолжается до первого безошибочного воспроизведения. Измеряется: 1) общее время заучивания, 2) число проб, 3) число правильных ответов в каждой пробе, 4) число ошибок в каждой пробе.
Процедура продолжается до первого безошибочного воспроизведения. Измеряется: 1) общее время заучивания, 2) число проб, 3) число правильных ответов в каждой пробе, 4) число ошибок в каждой пробе.
г) Метод сбережения. Суть метода в том, что испытуемый после первоначального заучивания и некоторого периода времени осуществляет повторное заучивание, которое должно удовлетворять следующим условиям: 1) должен применяться тот же метод, что и при первом заучивании; 2) необходимо использовать тот же критерий усвоения. Даже если по прошествии времени после первоначального заучивания испытуемый не может воспроизвести ни одного элемента материала, нельзя делать вывод о полном забывании, не применив метода повторного заучивания: если во второй раз ему понадобится меньше проб, чем в первый, то это значит, что какая-то часть материала в памяти все-таки сохранилась, но порог воспроизведения слишком высок (порог узнавания обычно значительно ниже). Измеряют:
Еа – число проб при первоначальном заучивании;
Еr – число проб при повторном заучивании;
J – число проб, соответствующее критерию усвоения (для первого безошибочного воспроизведения J=1).
Подсчитывают:
Абсолютное сбережение (Еа – Еr) – разность проб между первоначальным и повторным заучиванием;
Относительное сбережение (в процентах) по формуле:
Относительное сбережение необходимо подсчитывать для сравнения показателей нескольких испытуемых. Например, если испытуемый а заучил ряд за Еа=20 проб и Еr=16 проб, а испытуемый б за Еа=10 и Еr=6 проб, то абсолютное сбережение у обоих одинаково и равно Еа–Еr=4, в то время как относительное сбережение равно 20% и 40% соответственно, а это означает, что процесс запоминания у а в два раза продуктивней, чем у б.
д) Метод угадывания. Предложен Г. Мюллером и А. Пильцекером, психологами-ассоцианистами, и рекомендован Г. Эббингаузом к использованию. Ряд элементов предъявляется несколько раз, затем через определенное время экспериментатор воспроизводит этот ряд с пропусками некоторых элементов и предлагает угадать пропущенные элементы. Подсчитывают количество правильных ответов и ошибок.
Подсчитывают количество правильных ответов и ошибок.
Каждый из перечисленных методов имеет свои недостатки и преимущества (см. [3], с. 246-252), но поскольку условия и цели экспериментов неодинаковы, то использование того или иного метода зависит от обстоятельств.
3.3. Образование, сохранение и исчезновение ассоциаций
Важнейшим фактором, влияющим на образование и сохранение ассоциаций, является повторение. Чем чаще переживаются впечатления, образующие ассоциацию, тем с большей точностью и уверенностью они воспроизводятся и тем дольше сохраняются в памяти. Однако общее правило для оптимального количества повторений сформулировать крайне трудно: простые, но яркие события могут удерживаться много лет даже после однократного появления; события менее интересные и более сложные могут не остаться в памяти даже после многократного переживания. Для простейших случаев такие правила установлены (правило Йоста, см. раздел 4).
Кроме того, имеет значение принадлежность элементов запоминаемого ряда к единому целому. Если впечатления, образующие ассоциативную связь, организованы посредством ритма или рифмы, то они заучиваются быстрее. Так, стихотворная строфа запоминается гораздо легче, чем такое же количество не связанных между собой слов или прозаического текста, который, хотя и содержит смысловые связи, но не организован ритмически.
Если впечатления, образующие ассоциативную связь, организованы посредством ритма или рифмы, то они заучиваются быстрее. Так, стихотворная строфа запоминается гораздо легче, чем такое же количество не связанных между собой слов или прозаического текста, который, хотя и содержит смысловые связи, но не организован ритмически.
Также на образование ассоциативной связи влияют внимание и интерес. Вклад внимания часто не может быть возмещен никаким числом повторений, сколь много бы их ни было. Что касается эмоций, то “ассоциирующая сила удовольствия должна быть признана значительно большей, чем неудовольствия” ([3], с. 20). Ассоциативные связи, образующиеся благодаря чувству неудовольствия, тоже быстро доходят до сознания и могут существовать длительное время, однако “мысли человеческие имеют с определенной точки зрения возможность выбора, они предпочитают направление, ведущее к приятному. Возможность различных путей всегда им дана только прежним опытом и создавшимися на его основе ассоциациями, но какой путь они изберут, определяется, при прочих равных условиях, большей приятностью отдельных путей” ([3], с. 21).
21).
Ещё посмотрите лекцию «7 Расогенез» по этой теме.
С течением времени ассоциативная связь претерпевает изменения. Меняются представления, ее образующие, образы воспоминаний становятся смутными и неопределенными, связь между ними ослабевает, взаимное воспроизведение членов связи уже не происходит с прежней быстротой и точностью и может вовсе прекратиться.
Экспериментальные исследования Эббингауза, посвященные изменению памяти во времени, открыли новую эпоху в развитии психологии, а кривая забывания, получившая название кривой Эббингауза, приобрела значение методического образца, по которому в дальнейшем строились экспериментальные кривые. Материалом для исследования послужили специально изобретенные им бессмысленные слоги – сочетания двух согласных и гласной между ними, образованные так, чтобы они не вызывали смысловых ассоциаций. Г. Эббингауз придумал 2300 таких слогов, составлял из них ряды, которые затем заучивал сам или предлагал заучивать испытуемым. С помощью метода сбережений по результатам эксперимента была построена кривая, приведенная на рис. 2. Эта кривая отражает изменения, которые происходят в ассоциационной связи со временем – сначала она круто снижается, что соответствует быстрому забыванию заученного, затем ее падение замедляется и останавливается по истечении месяца на уровне 20% – сбереженный материал.
С помощью метода сбережений по результатам эксперимента была построена кривая, приведенная на рис. 2. Эта кривая отражает изменения, которые происходят в ассоциационной связи со временем – сначала она круто снижается, что соответствует быстрому забыванию заученного, затем ее падение замедляется и останавливается по истечении месяца на уровне 20% – сбереженный материал.
Главной целью Эббингауза было нахождение “чистых” законов памяти, которые не зависели бы от индивидуальных различий, установок испытуемых и экспериментаторов, т.е. носили бы всеобщий характер. Именно поэтому изобретение бессмысленных слогов обеспечило ему успех, поскольку категория значения слова была устранена и отпала необходимость пользоваться методом интроспекции. Э. Титченер, ученик В. Вундта, назвал изобретение бессмысленных слогов наиболее важным событием в психологии со времен Аристотеля. Очищенные от смысла, они позволяли проникнуть в область высших психических функций, вычленив общий для всех момент научения и усвоения.
3.4. Критика ассоциативной теории
Несмотря на достижения в области экспериментального исследования памяти, ассоциативная теория памяти обладала рядом недостатков. Ассоцианизм не различал общего и специфического, прямо отождествив их, и поэтому при каждом новом столкновении со специфическим данная концепция оказывалась несостоятельной. Неоднократно отмечалось, что даже при использовании бессмысленных слогов нельзя до конца избавиться от “искажающего” влияния смысла: испытуемым все-таки удавалось ассоциировать эти слоги с другими словами и образами, что облегчало запоминание. Оказалось, что реальная работа памяти не сводится к установлению ассоциаций, имеют место также и другие механизмы запоминания, что особенно ярко проявляется в работе высших форм памяти. Запоминание в экспериментах Эббингауза происходило в искусственно созданных лабораторных условиях, и поэтому в стороне от исследования оставались такие факторы, как установки, мотивы и цели мнестической деятельности и т. д. Однако принцип ассоциации до сих пор используется в психологии и не утратил своего значения: например, в современных когнитивных теориях памяти (см. раздел 6) он рассматривается как основной принцип устройства долговременной памяти.
д. Однако принцип ассоциации до сих пор используется в психологии и не утратил своего значения: например, в современных когнитивных теориях памяти (см. раздел 6) он рассматривается как основной принцип устройства долговременной памяти.
Подробнее см. работы [1; 3; 18, с. 249-261]
Ассоциативная симметрия и теория памяти
Андерсон, Дж. А., Сильверстайн, Дж. В., Ритц, С. А., и Джонс, Р. С. (1977). Отличительные черты, категориальное восприятие и вероятностное обучение: некоторые приложения нейронной модели. Психологический обзор , 84 , 413–451.
Артикул Google Scholar
Аш, С.Э. (1968). Доктринальная тирания ассоциативизма: или что плохого в зубрежке. В Т. Р. Диксон и Д. Л. Хортон (ред.), Вербальное поведение и общая теория поведения (стр. 214–228). Энглвуд Клиффс, Нью-Джерси: Прентис-Холл.
Google Scholar
Аш, С.
 Э., и Эбенхольц, С.М. (1962). Принцип ассоциативной симметрии. Труды Американского философского общества , 106 , 135–163.
Э., и Эбенхольц, С.М. (1962). Принцип ассоциативной симметрии. Труды Американского философского общества , 106 , 135–163.Google Scholar
Бартлинг, К. А., и Томпсон, П. (1977). Специфичность кодирования: асимметрия поиска в парадигме отказа распознавания. Журнал экспериментальной психологии: обучение и память человека , 3 , 690–700.
Артикул Google Scholar
Бегг, И. (1979). Потеря трассировки и сбой распознавания неназванных слов. Память и познание , 7 , 113–123.
Артикул Google Scholar
Бишоп, Ю.М.М., Финберг, С.Е., и Холланд, П.В. (1975). Дискретный многомерный анализ: теория и практика . Кембридж, Массачусетс: MIT Press.
Google Scholar
Браун, Д. А., Даллоз, П., и Хьюм, К.
 (1995). Математические и коннекционистские модели памяти человека: сравнение. Память , 2 , 113–145.
(1995). Математические и коннекционистские модели памяти человека: сравнение. Память , 2 , 113–145.Артикул Google Scholar
Браун, Т. Х., и Чаттарджи, С. (1995). Синаптическая пластичность Хебба. В MA Arbib (Ed.), . Справочник по теории мозга и нейронным сетям (стр. 454–459). Кембридж, Массачусетс: MIT Press.
Google Scholar
Бухманн Дж., Дивко Р. и Шультен К. (1989). Ассоциативная память с высокой информативностью. Physical Review A , 39 , 2689–2692.
Артикул пабмед Google Scholar
Чаппелл, М., и Хамфрис, М.С. (1994). Автоассоциативная нейронная сеть для разреженных представлений: анализ и применение к моделям распознавания и отзыва по сигналу. Психологический обзор , 101 , 103–128.
Артикул Google Scholar (1958).
 Взаимодействие способности и количества практики со значимостью стимула и реакции (m, m’) в парно-ассоциированном обучении. Журнал экспериментальной психологии , 56 , 193–202.
Взаимодействие способности и количества практики со значимостью стимула и реакции (m, m’) в парно-ассоциированном обучении. Журнал экспериментальной психологии , 56 , 193–202.Артикул пабмед Google Scholar
Доннелли, Р. Э. (1988). Эффекты прайминга в последовательных эпизодических тестах. Журнал экспериментальной психологии: обучение, память и познание , 14 , 256–265.
Артикул Google Scholar
Эббингауз, Х. (1885/1913). Память: вклад в экспериментальную психологию (HA Ruge & CA Bussenius, Trans.). Нью-Йорк: Педагогический колледж Колумбийского университета. (Оригинальная работа опубликована в 1885 г.)
Google Scholar
Экстранд, Б. Р. (1966). Обратные ассоциации. Психологический бюллетень , 65 , 50–64.
Артикул пабмед Google Scholar
Эстес, В.
 К. (1960). Теория обучения и новая «ментальная химия». Психологический обзор , 67 , 207–223.
К. (1960). Теория обучения и новая «ментальная химия». Психологический обзор , 67 , 207–223.Артикул пабмед Google Scholar
Флекссер, А. Дж. (1981). Гомогенизация таблицы непредвиденных обстоятельств 2 3 2: метод устранения зависимостей из-за различий в предметах и элементах. Психологический обзор , 88 , 327–339.
Артикул Google Scholar
Френдли, М., Франклин, П.Е., Хоффман, Д., и Рубин, Д.К. (1982). Пул слов Торонто: нормы образности, конкретности, орфографических переменных и грамматического использования для 1080 слов. Методы и приборы исследования поведения , 14 , 375–399.
Артикул Google Scholar
Гардинер, Дж. М., Грегг, В. Х., и Хэмптон, Дж. А. (1988). Частота слов и эффекты генерации. Журнал экспериментальной психологии: обучение, память и познание , 14 , 687–693.

Артикул Google Scholar
Гиллунд Г. и Шиффрин Р. М. (1984). Поисковая модель как для узнавания, так и для припоминания. Психологический обзор , 91 , 1–67.
Артикул пабмед Google Scholar
Грино, Дж. Г., Джеймс, К. Т., и ДаПолито, Ф. Дж. (1971). Когнитивная интерпретация отрицательного переноса и забывания парных партнеров. Журнал вербального обучения и вербального поведения , 10 , 331–345.
Артикул Google Scholar
Герц, Дж., Крог, А., и Палмер, Р. Г. (1991). Введение в теорию нейронных вычислений . Редвуд-Сити, Калифорния: Аддисон-Уэсли.
Google Scholar
Хинцман, Д. (1987). Узнавание и отзыв в MINERVA 2: анализ парадигмы «узнавание-неудача». В П. Моррисе (ред.), Моделирование познания (стр.
 215–229). Нью-Йорк: Уайли.
215–229). Нью-Йорк: Уайли.Google Scholar
Хопфилд, Дж. Дж. (1982). Нейронные сети и физические системы с возникающими коллективными вычислительными способностями. Труды Национальной академии наук , 84 , 8429–8433.
Артикул Google Scholar
Горовиц, Л.М., Браун, З.М., и Вайсблут, С. (1964). Наличие и направленность ассоциаций. Журнал экспериментальной психологии , 68 , 541–549.
Артикул пабмед Google Scholar
Горовиц, Л. М., Норман, С. А., и Дэй, Р. С. (1966). Доступность и ассоциативная симметрия. Психологический обзор , 73 , 1–15.
Артикул пабмед Google Scholar
Ховард, М. В., и Кахана, М. Дж. (1999). Эффекты контекстуальной изменчивости и последовательного положения в свободном воспоминании.
 Журнал экспериментальной психологии: обучение, память и познание , 25 , 923–941.
Журнал экспериментальной психологии: обучение, память и познание , 25 , 923–941.Артикул Google Scholar
Ховард, М. В., и Кахана, М. Дж. (2002). Распределенное представление временного контекста. Журнал математической психологии , 46 , 269–299.
Артикул Google Scholar
Хамфрис, М.С., Бейн, Дж.Д., и Пайк, Р. (1989). Различные способы подсказки когерентной системы памяти: теория эпизодических, семантических и процедурных задач. Психологический обзор , 96 , 208–233.
Артикул Google Scholar
Хамфрис, М. С., и Бойер, П. А. (1980). Эффекты последовательного тестирования и взаимосвязь между распознаванием и неудачным распознаванием. Память и познание , 8 , 271–277.
Артикул Google Scholar
Хамфрис, М.
 С., Пайк, Р., Бейн, Дж.Д., и Техан, Г. (1989). Глобальное сопоставление: сравнение моделей SAM, Minerva II, Matrix и TODAM. Журнал математической психологии , 33 , 36–67.
С., Пайк, Р., Бейн, Дж.Д., и Техан, Г. (1989). Глобальное сопоставление: сравнение моделей SAM, Minerva II, Matrix и TODAM. Журнал математической психологии , 33 , 36–67.Артикул Google Scholar
Джордан, Мичиган (1986). Введение в линейную алгебру в параллельной распределенной обработке. В DE Rumelhart & JL McClelland (Eds.), Параллельная распределенная обработка: исследование микроструктуры познания. Том 1: Foundations (стр. 365–422). Кембридж, Массачусетс: MIT Press.
Google Scholar
Кахана, М.Дж. (1996). Ассоциативные поисковые процессы в свободном воспоминании. Память и познание , 24 , 103–109.
Артикул Google Scholar
Кахана, М.Дж. (2000). Случайный анализ памяти. В E. Tulving & FIM Craik (Eds.), Оксфордский справочник по человеческой памяти (стр.
 323–384). Оксфорд: Издательство Оксфордского университета.
323–384). Оксфорд: Издательство Оксфордского университета.Google Scholar
Кахана, М.Дж., и Каплан, Дж.Б. (2002). Ассоциативная асимметрия в пробном воспроизведении серийных списков. Память и познание , 30 , 841–849.
Артикул Google Scholar
Кахана, М.Дж., Ховард, М.В., Заромб, Ф., и Вингфилд, А. (2002). Возраст разделяет эффекты давности и отставания в свободном воспоминании. Журнал экспериментальной психологии: обучение, память и познание , 28 , 530–540.
Артикул Google Scholar
Кахана, М.Дж., Риццуто, Д.С., и Шнайдер, А. (2002). Анализ отношения распознавания-припоминания в четырех моделях распределенной памяти . Рукопись представлена для публикации.
Келер, В. (1947). Гештальт-психология . Нью-Йорк: Ливерит.
Google Scholar
Леви, К.
 М., и Невилл, Д.Д. (1974). B-A обучение как функция степени A-B обучения. Журнал экспериментальной психологии , 102 , 327–329.
М., и Невилл, Д.Д. (1974). B-A обучение как функция степени A-B обучения. Журнал экспериментальной психологии , 102 , 327–329.Артикул Google Scholar
Локхарт, Р. С. (1969). Восстановительная асимметрия в воспоминании прилагательных и существительных. Журнал экспериментальной психологии , 79 , 12–17.
Артикул Google Scholar
Мандлер, Г. (1959). Переменные стимула и предметные переменные: предостережение. Психологический обзор , 66 , 145–149.
Артикул пабмед Google Scholar
Мандлер, Г., Рабинович, Дж. К., и Саймон, Р. А. (1981). Координатная организация: целостное представление пар слов. Американский журнал психологии , 92 , 209–222.
Артикул Google Scholar
Мартин Э.
 (1971). Теория вербального обучения и явления независимого поиска. Психологический обзор , 78 , 314–332.
(1971). Теория вербального обучения и явления независимого поиска. Психологический обзор , 78 , 314–332.Артикул Google Scholar
Мартин, Э., и Грино, Дж. Г. (1972). Независимость ассоциаций проверена. Психологический обзор , 79 , 265–267.
Артикул Google Scholar
Макнотон, Б.Л., и Моррис, Р.Г. (1987). Синаптическое усиление гиппокампа и хранение информации в распределенной системе памяти. Тенденции в нейронауках , 10 , 408–415.
Артикул Google Scholar
Меткалф, Дж. (1985). Уровни обработки, специфичность кодирования, проработка и CHARM. Психологический обзор , 92 , 1–38.
Артикул Google Scholar
Меткалф, Дж. (1991). Ошибка распознавания и составная трассировка памяти в CHARM.
 Психологический обзор , 98 , 529–553.
Психологический обзор , 98 , 529–553.Артикул пабмед Google Scholar
Меткалф, Дж. (1993). Мониторинг новизны, метапознание и контроль в сложной голографической модели ассоциативного воспоминания: последствия для амнезии Корсакова. Психологический обзор , 100 , 3–22.
Артикул пабмед Google Scholar
Мердок, Б. Б. (1956). Обратное обучение в парных ассоциатах. Журнал экспериментальной психологии , 51 , 213–215.
Артикул пабмед Google Scholar
Мердок, Б. Б. (1962). Эффект последовательного положения свободного припоминания. Журнал экспериментальной психологии , 64 , 482–488.
Артикул Google Scholar
Мердок, Б. Б. (1965). Ассоциативная симметрия и дихотическое представление.
 Журнал вербального обучения и вербального поведения , 4 , 222–226.
Журнал вербального обучения и вербального поведения , 4 , 222–226.Артикул Google Scholar
Мердок, Б. Б. (1966). Прямые и обратные ассоциации в парных ассоциатах. Журнал экспериментальной психологии , 71 , 732–737.
Артикул пабмед Google Scholar
Мердок, Б. Б. (1979). Свертка и корреляция в восприятии и памяти. В Л.-Г. Нильссон (ред.), Перспективы исследования памяти: очерки в честь 500-летия Упсальского университета (стр. 105–119). Хиллсдейл, Нью-Джерси: Эрлбаум.
Google Scholar
Мердок, Б. Б. (1982). Теория хранения и поиска элементов и ассоциативной информации. Психологический обзор , 89 , 609–626.
Артикул Google Scholar
Мердок, Б. Б. (1985). Сверточные и матричные системы: ответ Пайку.
 Психологический обзор , 92 , 130–132.
Психологический обзор , 92 , 130–132.Артикул Google Scholar
Мердок, Б. Б. (1989). Обучение в модели распределенной памяти. In C. Izawa (Ed.), Текущие проблемы когнитивных процессов: Симпозиум Floweree по познанию (стр. 69–106). Хиллсдейл, Нью-Джерси: Эрлбаум.
Google Scholar
Мердок, Б. Б. (1995). Сходство в модели распределенной памяти. Журнал математической психологии, 39 , 251–264.
Артикул Google Scholar
Мердок, Б. Б. (1997). Контекст и посредники в теории распределенной ассоциативной памяти (TODAM2). Психологический обзор , 104 , 839–862.
Артикул Google Scholar
Нэрн, Дж. С., и Виднер, Р. Л. (1988). Знакомство и лексика как детерминанты эффекта генерации. Журнал экспериментальной психологии: обучение, память и познание , 14 , 694–699.

Артикул Google Scholar
Нильссон, Л.-Г., и Гардинер, Дж. М. (1993). Выявление исключений в базе данных исследований нарушений распознавания с 1973 по 1992 год. Память и познание , 21 , 397–410.
Артикул Google Scholar
Пайк, Р. (1984). Сравнение систем свертки и матричной распределенной памяти для ассоциативного припоминания и распознавания. Психологический обзор , 91 , 281–294.
Артикул Google Scholar
Примофф, Э. (1938). Обратные и прямые ассоциации как организующее действие в последовательном и парно-ассоциированном обучении. Журнал психологии , 5 , 375–395.
Артикул Google Scholar
Raaijmakers, JGW, &Shiffrin, R.M. (1980). СЭМ: Теория вероятностного поиска ассоциативной памяти.
 В GH Bower (Ed.), Психология обучения и мотивации: достижения в исследованиях и теории (Том 14, стр. 207–262). Нью-Йорк: Академическая пресса.
В GH Bower (Ed.), Психология обучения и мотивации: достижения в исследованиях и теории (Том 14, стр. 207–262). Нью-Йорк: Академическая пресса.Google Scholar
Риццуто, Д.С., и Кахана, М.Дж. (2001). Автоассоциативная модель парного ассоциированного обучения. Нейронные вычисления , 13 , 2075–2092.
Артикул пабмед Google Scholar
Робинсон, Э. С. (1932). Теория ассоциаций сегодня; эссе по систематической психологии . Нью-Йорк: Век.
Книга Google Scholar
Рок, И. (1957). Роль повторения в ассоциативном обучении. Американский журнал психологии , 70 , 186–193.
Артикул пабмед Google Scholar
Рок, И., и Серасо, Дж. (1964). К когнитивной теории ассоциативного обучения.
 В C. Scheerer (Ed.), Познание: теория, исследования и перспективы (стр. 110–146). Нью-Йорк: Харпер и Роу.
В C. Scheerer (Ed.), Познание: теория, исследования и перспективы (стр. 110–146). Нью-Йорк: Харпер и Роу.Google Scholar
Сламецка, Нью-Джерси (1976). Анализ списков двойной функции. Память и познание , 4 , 581–585.
Артикул Google Scholar
Тревес, А., и Роллс, Э. Т. (1994). Вычислительный анализ роли гиппокампа в памяти. Гиппокамп , 4 , 374–391.
Артикул пабмед Google Scholar
Тульвинг, Э. (1964). Удержание во время и между испытаниями: примечания к теории вербального обучения со свободным отзывом. Психологический обзор , 71 , 219–237.
Артикул пабмед Google Scholar
Тульвинг, Э. (1983). Элементы эпизодической памяти . Нью-Йорк: Издательство Оксфордского университета.

Google Scholar
Тульвинг, Э., и Арбакл, Т.Ю. (1966). Входные и выходные помехи в кратковременной ассоциативной памяти. Журнал экспериментальной психологии , 72 , 145–150.
Артикул пабмед Google Scholar
Тульвинг, Э., и Томпсон, Д.М. (1973). Специфика кодирования и поисковые процессы в эпизодической памяти. Психологический обзор , 80 , 352–373.
Артикул Google Scholar
Тульвинг, Э., и Уайзман, С. (1975). Связь между узнаванием и отказом узнавания запоминаемых слов. Бюллетень Психономического общества , 6 , 79–82.
Google Scholar
Уорд Г. и Вудворд Г. (2002). Три фактора, вызывающие эффект частотности слов при свободном вспоминании . Рукопись представлена для публикации.

Во, Северная Каролина (1970). Ассоциативная симметрия и задержки припоминания: различие между обучением и производительностью. Acta Psychologica , 33 , 326–337.
Артикул Google Scholar
Weber, EU (1988). Ожидание и дисперсия распределений сходства элементов в сверточно-корреляционной модели распределенной памяти. Журнал математической психологии , 32 , 1–43.
Артикул Google Scholar
Вулфорд, Г. (1971). Функция отдельных ассоциаций для парно-ассоциативной производительности. Психологический обзор , 78 , 303–313.
Артикул Google Scholar
Воллен, К.А., Фокс, Р.А., и Лоури, Д.Х. (1970). Вариации асимметрии в зависимости от степени опережающего обучения. Журнал вербального обучения и вербального поведения , 86 , 416–419.

Google Scholar
Скачать ссылки
Ассоциативное обучение и гиппокамп
Ассоциативная память определяется как способность узнавать и запоминать отношения между не связанными друг с другом элементами, такими как имя кого-то, кого мы только что встретили, или аромат определенных духов.
Венди А. Судзуки, доктор философии
Каждый день мы узнаем и запоминаем множество новой информации от имен новых людей, которых мы встречаем, до лучшего блюда, которое можно заказать в том или ином ресторане, до расположения новой велосипедной дорожки. Эта новая информация сначала приобретается, и, если она усиливается в процессе, называемом консолидацией, в конечном итоге сохраняется в долговременной памяти. Структуры медиальной височной доли необходимы для этой способности приобретать новые долговременные воспоминания о фактах и событиях. Эта форма памяти называется декларативной памятью у людей и реляционной памятью у животных. Конвергентные результаты нейропсихологических исследований у людей, а также экспериментальные исследования повреждений и нейроанатомические исследования на животных показали, что ключевые структуры медиальной височной доли, важные для декларативной/реляционной памяти, включают гиппокамп вместе с окружающей энторинальной, периринальной и парагиппокампальной корой. Хотя ясно, что структуры медиальной височной доли необходимы для приобретения новых декларативных/реляционных воспоминаний, у нас все еще есть лишь рудиментарное понимание нормальных паттернов нейронной активности, лежащих в основе этой способности. Чтобы начать рассмотрение этого вопроса, моя лаборатория зарегистрировала активность отдельных нейронов в гиппокампе, когда обезьяны выполняют различные задачи, требующие памяти. Мы сосредоточились на одной конкретной форме декларативной/реляционной памяти, называемой ассоциативной памятью, которая определяется как способность изучать и запоминать отношения между не связанными между собой элементами, такими как имя кого-то, кого мы только что встретили, или аромат определенных духов.
Конвергентные результаты нейропсихологических исследований у людей, а также экспериментальные исследования повреждений и нейроанатомические исследования на животных показали, что ключевые структуры медиальной височной доли, важные для декларативной/реляционной памяти, включают гиппокамп вместе с окружающей энторинальной, периринальной и парагиппокампальной корой. Хотя ясно, что структуры медиальной височной доли необходимы для приобретения новых декларативных/реляционных воспоминаний, у нас все еще есть лишь рудиментарное понимание нормальных паттернов нейронной активности, лежащих в основе этой способности. Чтобы начать рассмотрение этого вопроса, моя лаборатория зарегистрировала активность отдельных нейронов в гиппокампе, когда обезьяны выполняют различные задачи, требующие памяти. Мы сосредоточились на одной конкретной форме декларативной/реляционной памяти, называемой ассоциативной памятью, которая определяется как способность изучать и запоминать отношения между не связанными между собой элементами, такими как имя кого-то, кого мы только что встретили, или аромат определенных духов. В частности, мы исследовали паттерны активности нейронов гиппокампа, когда обезьяны находятся в процессе формирования новых ассоциаций в памяти. Мы предположили, что если гиппокамп важен для раннего формирования новых ассоциаций, мы должны увидеть изменения в нейронной активности, которые параллельны поведенческому обучению. Наша долгосрочная цель состоит в том, чтобы понять эволюцию сигналов, связанных с обучением, по всей медиальной височной доле, поскольку воспоминания изначально устанавливаются, укрепляются и в конечном итоге сохраняются в долговременной памяти.
В частности, мы исследовали паттерны активности нейронов гиппокампа, когда обезьяны находятся в процессе формирования новых ассоциаций в памяти. Мы предположили, что если гиппокамп важен для раннего формирования новых ассоциаций, мы должны увидеть изменения в нейронной активности, которые параллельны поведенческому обучению. Наша долгосрочная цель состоит в том, чтобы понять эволюцию сигналов, связанных с обучением, по всей медиальной височной доле, поскольку воспоминания изначально устанавливаются, укрепляются и в конечном итоге сохраняются в долговременной памяти.
Задача, требующая нового ассоциативного обучения
Чтобы изучить закономерности нейронной активности во время формирования ассоциативной памяти, мы обучили двух обезьян выполнять задание на ассоциацию места и сцены. В этом задании от животных требовалось выучить новые ассоциации между конкретными сложными визуальными «сценами» и конкретными целевыми местами с вознаграждением. Мы знаем, что медиальная височная доля играет важную роль в нормальном выполнении этой задачи, поскольку повреждение этой области у обезьян приводит к значительному ухудшению способности запоминать новые ассоциации место-сцена (Brasted et al. , 2003; Brasted et al., 2002). ; Мюррей и др., 2000; Уайз и Мюррей, 19 лет.99; Мюррей и Уайз, 1996 г.; Рупняк и Гаффан, 1987). Схематическое изображение задачи показано на рисунке 1 . В каждом испытании обезьянам сначала показывают 4 идентичных целевых стимула, наложенных на сложную визуальную сцену (обычно изображение реальной сцены на открытом воздухе). После интервала задержки, в течение которого сцена исчезает, но цели остаются на экране, животному дают сигнал сделать одно движение глаз к одной из четырех периферийных целей на экране. Для каждой визуальной сцены только одна из 4 целей связана с наградой в виде сока. Каждый день животные методом проб и ошибок усваивали 2-4 новые ассоциации место-сцена. Новые ассоциации «место-сцена» случайным образом смешивались с хорошо изученными «эталонной» ассоциацией, которую животные видели за много месяцев до начала экспериментов по записи. Каждая из 4 эталонных сцен была связана с различным целевым местоположением (т.
, 2003; Brasted et al., 2002). ; Мюррей и др., 2000; Уайз и Мюррей, 19 лет.99; Мюррей и Уайз, 1996 г.; Рупняк и Гаффан, 1987). Схематическое изображение задачи показано на рисунке 1 . В каждом испытании обезьянам сначала показывают 4 идентичных целевых стимула, наложенных на сложную визуальную сцену (обычно изображение реальной сцены на открытом воздухе). После интервала задержки, в течение которого сцена исчезает, но цели остаются на экране, животному дают сигнал сделать одно движение глаз к одной из четырех периферийных целей на экране. Для каждой визуальной сцены только одна из 4 целей связана с наградой в виде сока. Каждый день животные методом проб и ошибок усваивали 2-4 новые ассоциации место-сцена. Новые ассоциации «место-сцена» случайным образом смешивались с хорошо изученными «эталонной» ассоциацией, которую животные видели за много месяцев до начала экспериментов по записи. Каждая из 4 эталонных сцен была связана с различным целевым местоположением (т. е. с севером, югом, востоком или западом). Реакция на эталонные сцены использовалась для контроля возможной двигательной активности в клетках гиппокампа.
е. с севером, югом, востоком или западом). Реакция на эталонные сцены использовалась для контроля возможной двигательной активности в клетках гиппокампа.
Паттерны активности гиппокампа при изучении новых ассоциаций место-сцена
В наших первоначальных исследованиях мы сосредоточились на гиппокампе, медиальной структуре височной доли, долгое время участвующей в ассоциативном обучении и памяти (Eichenbaum and Cohen, 2001; Squire and Zola , 1996; Сковилл и Милнер, 1957). Сначала мы спросили, реагируют ли клетки гиппокампа по-разному на различные визуальные сцены, используемые в задаче. Мы обнаружили, что 61% изолированных клеток гиппокампа были вовлечены в задачу, поскольку они по-разному реагировали на разные сцены (то есть визуально избирательный ответ). Более того, в соответствии с нашей рабочей гипотезой мы обнаружили, что 28% избирательно реагирующих клеток (18% всей зарегистрированной популяции клеток гиппокампа) демонстрировали изменения в нервной активности в ходе испытаний, которые в значительной степени коррелировали с поведенческой кривой обучения животного для конкретного исследования. сцена. Мы назвали эти клетки «изменяющимися клетками». Наблюдались две категории изменяющихся клеток. Устойчивые изменяющиеся клетки (54% популяции изменяющихся клеток) сигнализировали об обучении изменением нейронной активности, которое сохранялось до тех пор, пока мы могли удерживать клетку (обычно от 30 минут до 1 часа). Многие из этих клеток демонстрировали резкое увеличение нейронной активности, которое соответствовало кривой поведенческого обучения животных для этой связи (рис. 2А-9).0758). Важно отметить, что эти обучающие сигналы были очень избирательными в том смысле, что меняющаяся клетка обычно меняет свою активность только для одной конкретной изученной сцены, в то время как ответы на другие изученные сцены не меняются с течением времени. Одна из интерпретаций результатов, показанных на рис. 2A , заключается в том, что изменение нейронной активности связано с обучением. Однако другая возможная интерпретация заключается в том, что эта деятельность связана с обучением определенной двигательной реакции (т.
сцена. Мы назвали эти клетки «изменяющимися клетками». Наблюдались две категории изменяющихся клеток. Устойчивые изменяющиеся клетки (54% популяции изменяющихся клеток) сигнализировали об обучении изменением нейронной активности, которое сохранялось до тех пор, пока мы могли удерживать клетку (обычно от 30 минут до 1 часа). Многие из этих клеток демонстрировали резкое увеличение нейронной активности, которое соответствовало кривой поведенческого обучения животных для этой связи (рис. 2А-9).0758). Важно отметить, что эти обучающие сигналы были очень избирательными в том смысле, что меняющаяся клетка обычно меняет свою активность только для одной конкретной изученной сцены, в то время как ответы на другие изученные сцены не меняются с течением времени. Одна из интерпретаций результатов, показанных на рис. 2A , заключается в том, что изменение нейронной активности связано с обучением. Однако другая возможная интерпретация заключается в том, что эта деятельность связана с обучением определенной двигательной реакции (т. Е. Обучением реагировать на север). В соответствии с этой интерпретацией ранние правильные пробы, возможно, не вызвали большой активности, если движения были сделаны предварительно, но сильная двигательная активность наблюдается, как только животное начинает последовательно реагировать на север. Если бы эта интерпретация, основанная на моторике, была верна, мы ожидали бы увидеть аналогичные уровни активности этой клетки в ответ на эталонную сцену с тем же целевым расположением на севере. Такого никогда не было. Фактически, меняющиеся клетки обычно реагировали практически без активности на эталонную сцену с тем же вознагражденным целевым местоположением. Эти результаты подтверждают идею о том, что изменение активности связано с изучением новой связи между сценой и движением цели/глаза, а не с изучением конкретной двигательной реакции. С этой идеей также согласуются результаты других контрольных экспериментов, в которых животные выучили 2 последовательных набора новых ассоциаций места и сцены.
Е. Обучением реагировать на север). В соответствии с этой интерпретацией ранние правильные пробы, возможно, не вызвали большой активности, если движения были сделаны предварительно, но сильная двигательная активность наблюдается, как только животное начинает последовательно реагировать на север. Если бы эта интерпретация, основанная на моторике, была верна, мы ожидали бы увидеть аналогичные уровни активности этой клетки в ответ на эталонную сцену с тем же целевым расположением на севере. Такого никогда не было. Фактически, меняющиеся клетки обычно реагировали практически без активности на эталонную сцену с тем же вознагражденным целевым местоположением. Эти результаты подтверждают идею о том, что изменение активности связано с изучением новой связи между сценой и движением цели/глаза, а не с изучением конкретной двигательной реакции. С этой идеей также согласуются результаты других контрольных экспериментов, в которых животные выучили 2 последовательных набора новых ассоциаций места и сцены. Мы обнаружили, что изменение клеток, идентифицированных в первом наборе изученных ассоциаций место-сцена, никогда не сигнализировало об обучении второму новому набору ассоциаций место-сцена, даже когда поощряемое целевое местоположение было тем же самым (т. Е. Вознаграждалась северная цель).
Мы обнаружили, что изменение клеток, идентифицированных в первом наборе изученных ассоциаций место-сцена, никогда не сигнализировало об обучении второму новому набору ассоциаций место-сцена, даже когда поощряемое целевое местоположение было тем же самым (т. Е. Вознаграждалась северная цель).
Остальные 45% изменяющихся клеток демонстрировали другой паттерн связанной с обучением активности. Эти изменяющиеся клетки начинались с избирательной реакции на сцену либо во время сцены, либо в период задержки задачи в начале сеанса, задолго до того, как животное научилось ассоциации. Эти клетки сигнализировали об обучении, возвращаясь к исходной активности, и это возвращение к исходному уровню, как правило, антикоррелировало с кривой обучения животного для этой конкретной сцены (рис. 2В, ). Мы назвали эти клетки базовыми устойчивыми изменяющимися клетками. Важно отметить, что изменения в нервной активности, наблюдаемые в исходно устойчивых клетках, были столь же избирательны для конкретной заученной сцены, как и в устойчивых изменяющихся клетках. Подобные паттерны активности никогда не наблюдались для эталонной сцены с соответствующим вознаграждаемым целевым местоположением, что позволяет предположить, что эти сигналы не были двигательными.
Подобные паттерны активности никогда не наблюдались для эталонной сцены с соответствующим вознаграждаемым целевым местоположением, что позволяет предположить, что эти сигналы не были двигательными.
Таким образом, как клетки с устойчивым изменением, так и клетки с устойчивым изменением базового уровня обеспечивают высокоселективный сигнал о том, когда конкретная сцена изучена. Мы предполагаем, что эти избирательные увеличения и уменьшения нейронной активности, которые происходят в популяции гиппокампа, могут представлять собой обучающий сигнал гиппокамповой сети. Будет важно изучить обучающие сигналы по большему количеству клеток гиппокампа, записанные одновременно, чтобы лучше понять взаимодействие между клетками во время обучения.
Синхронизация активности гиппокампа во время изучения новых ассоциаций место-сцена
Важным вопросом для любого исследования, изучающего нейронные корреляты поведения, является определение причинно-следственной связи между наблюдаемыми паттернами нейронной активности и поведенческим результатом.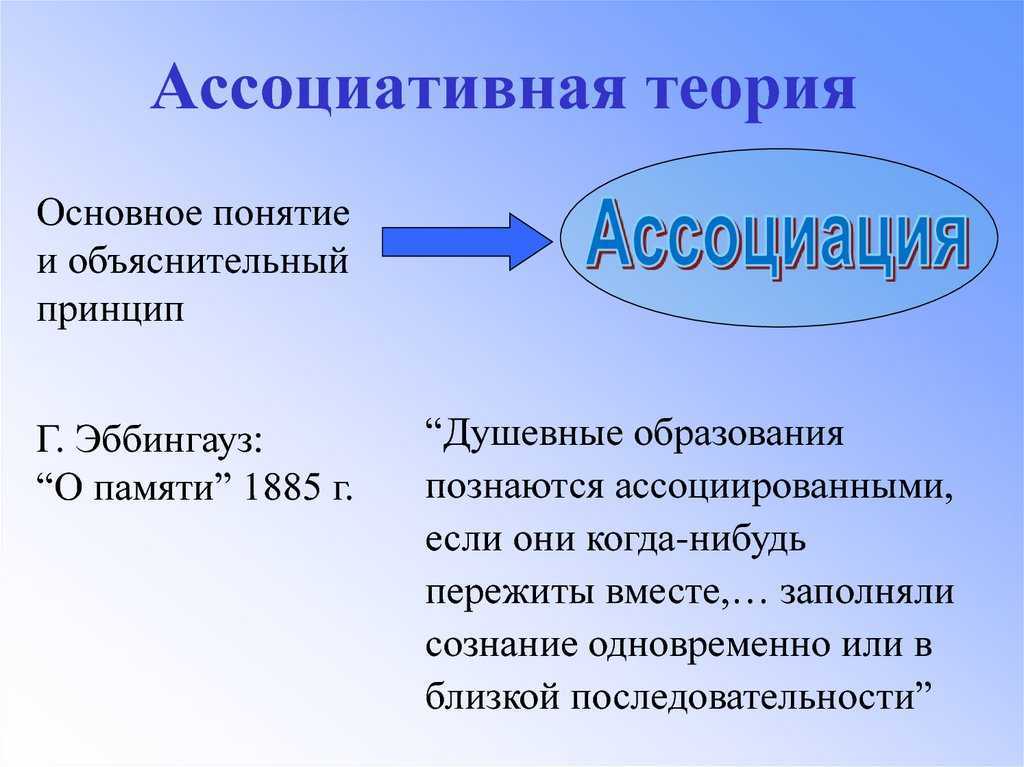 Является ли наблюдаемая нейронная активность движущей силой поведения или она происходит ниже по течению от критических мест происхождения? Для решения этой проблемы использовались различные подходы. Например, как упоминалось выше, исследования поражений могут вовлекать конкретную область мозга в нормальное выполнение задачи, хотя сами по себе исследования поражений не могут определить паттерны нейронной активности, лежащие в основе этой функции. Исследования с электрической стимуляцией использовались для проверки влияния прямой стимуляции на выбор животных при проблемах сенсорной дискриминации (Salzman et al., 19).92; Salzman et al., 1990), но этот подход не применялся в исследованиях функции гиппокампа. Третий метод, который использовался для исследования взаимосвязи между нейронной активностью и поведением, заключается в изучении точного времени изменений нейронной активности по отношению к поведенческому обучению. Мы предполагаем, что те избирательные нейронные изменения, которые происходят до проявления поведенческого обучения, могут быть вовлечены в изменение поведения, в то время как те, которые происходят после поведенческого обучения, могут играть роль в укреплении вновь сформированной ассоциации.
Является ли наблюдаемая нейронная активность движущей силой поведения или она происходит ниже по течению от критических мест происхождения? Для решения этой проблемы использовались различные подходы. Например, как упоминалось выше, исследования поражений могут вовлекать конкретную область мозга в нормальное выполнение задачи, хотя сами по себе исследования поражений не могут определить паттерны нейронной активности, лежащие в основе этой функции. Исследования с электрической стимуляцией использовались для проверки влияния прямой стимуляции на выбор животных при проблемах сенсорной дискриминации (Salzman et al., 19).92; Salzman et al., 1990), но этот подход не применялся в исследованиях функции гиппокампа. Третий метод, который использовался для исследования взаимосвязи между нейронной активностью и поведением, заключается в изучении точного времени изменений нейронной активности по отношению к поведенческому обучению. Мы предполагаем, что те избирательные нейронные изменения, которые происходят до проявления поведенческого обучения, могут быть вовлечены в изменение поведения, в то время как те, которые происходят после поведенческого обучения, могут играть роль в укреплении вновь сформированной ассоциации. Чтобы ответить на этот вопрос, для всех изменяющихся клеток мы рассчитали количество попыток обучения и сравнили его с рассчитанным количеством попыток нейронных изменений (см. Wirth et al., 2003 для подробного описания используемых поведенческих и нейронных алгоритмов). Мы обнаружили, что изменяющиеся клетки гиппокампа могут опережать (14 примеров) параллельно (4 примера) и отставать (19 примеров).примеры) поведенческое обучение (рис. 3 ). Эти данные свидетельствуют о том, что гиппокамп участвует во всех стадиях процесса обучения, от нескольких попыток до того, как проявляется поведенческое обучение, когда наблюдаемая активность может быть вовлечена в управление поведенческими изменениями, лежащими в основе обучения, до нескольких попыток после обучения, когда активность может быть задействована. в укреплении новообразованной ассоциации.
Чтобы ответить на этот вопрос, для всех изменяющихся клеток мы рассчитали количество попыток обучения и сравнили его с рассчитанным количеством попыток нейронных изменений (см. Wirth et al., 2003 для подробного описания используемых поведенческих и нейронных алгоритмов). Мы обнаружили, что изменяющиеся клетки гиппокампа могут опережать (14 примеров) параллельно (4 примера) и отставать (19 примеров).примеры) поведенческое обучение (рис. 3 ). Эти данные свидетельствуют о том, что гиппокамп участвует во всех стадиях процесса обучения, от нескольких попыток до того, как проявляется поведенческое обучение, когда наблюдаемая активность может быть вовлечена в управление поведенческими изменениями, лежащими в основе обучения, до нескольких попыток после обучения, когда активность может быть задействована. в укреплении новообразованной ассоциации.
Резюме и будущие направления
Мы показали, что клетки в гиппокампе обеспечивают сильные паттерны нейронной активности, связанные с обучением, которые участвуют в начальном формировании новых ассоциативных воспоминаний. Поскольку эти изменения могут происходить до, в то же время или после обучения, эти данные позволяют предположить, что во время формирования новых ассоциативных воспоминаний может происходить постепенное вовлечение сети нейронов гиппокампа. Предыдущие исследования показали, что помимо гиппокампа клетки в нескольких других областях мозга, включая префронтальную кору (Asaad et al., 1998), лобные области, связанные с моторикой (Brasted and Wise, 2004; Chen and Wise, 1995a; Chen and Wise, 1995b; Mitz et al., 1991) и стриатум (Brasted and Wise, 2004) демонстрируют схожие паттерны, связанные с обучением. активность при выполнении сходных ассоциативных учебных заданий. Важной долгосрочной целью будет понимание того, как все эти области мозга от гиппокампа до связанных с моторикой областей лобной доли и полосатого тела могут работать вместе, чтобы лежать в основе начального формирования, а также раннего усиления и консолидации нового ассоциативного обучения.
Поскольку эти изменения могут происходить до, в то же время или после обучения, эти данные позволяют предположить, что во время формирования новых ассоциативных воспоминаний может происходить постепенное вовлечение сети нейронов гиппокампа. Предыдущие исследования показали, что помимо гиппокампа клетки в нескольких других областях мозга, включая префронтальную кору (Asaad et al., 1998), лобные области, связанные с моторикой (Brasted and Wise, 2004; Chen and Wise, 1995a; Chen and Wise, 1995b; Mitz et al., 1991) и стриатум (Brasted and Wise, 2004) демонстрируют схожие паттерны, связанные с обучением. активность при выполнении сходных ассоциативных учебных заданий. Важной долгосрочной целью будет понимание того, как все эти области мозга от гиппокампа до связанных с моторикой областей лобной доли и полосатого тела могут работать вместе, чтобы лежать в основе начального формирования, а также раннего усиления и консолидации нового ассоциативного обучения.
Ссылки
Асаад В. Ф., Райнер Г., Миллер Э. К. (1998) Нейронная активность в префронтальной коре приматов во время ассоциативного обучения. Нейрон 21: 1399-1407.
Ф., Райнер Г., Миллер Э. К. (1998) Нейронная активность в префронтальной коре приматов во время ассоциативного обучения. Нейрон 21: 1399-1407.
Brasted P.J., Bussey TJ, Murray EA, Wise SP (2002) Перерезка Fornix ухудшает условное зрительно-моторное обучение в задачах, связанных с непространственно дифференцированными ответами. J Нейрофизиол 87: 631-633.
Брастед П.Дж., Басси Т.Дж., Мюррей Э.А., Уайз С.П. (2003) Роль системы гиппокампа в ассоциативном обучении за пределами пространственной области. Мозг 126: 1202-1223.
Брастед П.Дж., Уайз С.П. (2004) Сравнение связанной с обучением активности нейронов в дорсальной премоторной коре и стриатуме. Eur J Neurosci 19: 721-740.
Chen L.L., Wise S.P. (1995a) Активность нейронов в дополнительном поле глаза при приобретении условных глазодвигательных ассоциаций. J Нейрофиз 73: 1101-1121.
Chen L.L., Wise S.P. (1995b) Дополнительное поле глаза контрастирует с передним полем глаза при приобретении условных глазодвигательных ассоциаций. J Нейрофиз 73: 1122-1134.
J Нейрофиз 73: 1122-1134.
Эйхенбаум Х., Коэн Н. Дж. (2001) От кондиционирования к сознательному воспоминанию. Нью-Йорк: Издательство Оксфордского университета.
Митц А. Р., Годшалк М., Уайз С. П. (1991) Зависимая от обучения активность нейронов в премоторной коре: активность при приобретении условных двигательных ассоциаций. J Neurosci 11: 1855-1872.
Мюррей Э. А., Басси Т. Дж., Уайз С. П. (2000) Роль префронтальной коры в сети произвольного зрительно-моторного картирования. Exp Br Res 133: 114-129.
Мюррей Э. А., Уайз С. П. (1996) Роль гиппокампа и нижележащей коры, но не миндалевидного тела, в зрительно-моторном условном обучении у макак-резусов. Behav Neurosci 110: 1261-1270.
Рупняк Н.М., Гаффан Д. (1987) Гиппокамп обезьяны и обучение пространственно направленным движениям. J Neurosci 7: 2331-2337.
Salzman C.D., Britten K.H., Newsome W.T. (1990) Корковая микростимуляция влияет на перцептивные суждения о направлении движения. Природа 346: 174-177.
Salzman C.D., Murasugi C.M., Britten K.H., Newsome W.T. (1992)Микростимуляция в зрительной области MT: влияние на эффективность распознавания направления. J Neurosci 12: 2331-2355.
Scoville W.B., Milner B. (1957) Потеря недавней памяти после двустороннего поражения гиппокампа. J Neurol Neurosurg Psych 20: 11-21.
Сквайр Л. Р., Зола С. М. (1996) Структура и функции декларативных и недекларативных систем памяти. Proc Natl Acad Sci 93: 13515-13522.
Вирт С., Янике М., Фрэнк Л.М., Смит А.С., Браун Е.Н., Судзуки В.А. (2003)Одиночные нейроны в гиппокампе обезьяны и изучение новых ассоциаций. Наука 300: 1578-1581.
Уайз С.П., Мюррей Э.А. (1999) Роль системы гиппокампа в условном моторном обучении: сопоставление предшественников с действием. Гиппокамп 9: 101-117.
Об авторе
Венди А. Судзуки получила докторскую степень в области неврологии в Калифорнийском университете в Сан-Диего в 1993 году и работала над докторской диссертацией в Лаборатории нейропсихологии Национального института психического здоровья.
