ДИССОЦИИРУЕТ — Что такое ДИССОЦИИРУЕТ?
Слово состоит из 12 букв: первая д, вторая и, третья с, четвёртая с, пятая о, шестая ц, седьмая и, восьмая и, девятая р, десятая у, одиннадцатая е, последняя т,
Слово диссоциирует английскими буквами(транслитом) — dissotsiiret
- Буква д встречается 1 раз. Слова с 1 буквой д
- Буква и встречается 3 раза. Слова с 3 буквами и
- Буква с встречается 2 раза. Слова с 2 буквами с
- Буква о встречается 1 раз. Слова с 1 буквой о
- Буква ц встречается 1 раз. Слова с 1 буквой ц
- Буква р встречается 1 раз. Слова с 1 буквой р
- Буква у встречается 1 раз.

- Буква е встречается 1 раз. Слова с 1 буквой е
- Буква т встречается 1 раз. Слова с 1 буквой т

Диссоциированный сон
Диссоциированный сон — медикаментозный сон, прерываемый в течение суток несколькими небольшими периодами бодрствования. Диссоциированный используется в лечебных целях. Somnus dissociatus
glossary.ru
Сон диссоциированный (s. dissociatus) используемый в лечебных целях медикаментозный С., прерываемый в течение суток несколькими небольшими периодами бодрствования.
Большой медицинский словарь. — 2000
Симптом диссоциированного Я
Симптом диссоциированного Я — вариант нарушения самоосознавания, при котором собственное Я воспринимается раздробленным на два функционально активных фрагмента и более…
Жмуров В.А. Большой толковый словарь терминов по психиатрии
Симптом диссоциированного Я – вариант нарушения самоосознавания, при котором собственное Я воспринимается раздробленным на два функционально относительно автономных фрагмента…
vocabulary.ru
- Слова из слова «диссоциирует»
- Слова на букву «д»
- Слова, начинающиеся на «ди»
- Слова c буквой «т» на конце
- Слова c «ет» на конце
- Слова, начинающиеся на «дис»
- Слова, начинающиеся на «дисс»
- Слова, оканчивающиеся на «ует»
- Слова, заканчивающиеся на «рует»
- диссоциация
- диссоциировавший
- диссоциировать
- диссоциирует
- диссоциируют
- диссоциирующий
Электролитическая диссоциация.
 Электролиты — материалы для подготовки к ЕГЭ по Химии
Электролиты — материалы для подготовки к ЕГЭ по ХимииЭлектрический ток – направленное движение заряженных частиц – электронов или ионов.
Электролиты – это вещества, растворы или расплавы (в ЕГЭ чаще речь о растворах) которых проводят электрический ток, то есть содержат заряженные частицы. Свободных электронов в растворе не бывает, носителями заряда являются ионы. Электрический ток проводят расплавы веществ с ионной кристаллической решеткой.
К электролитам относятся:
- Соли
- Кислоты
- Основания
Чем больше в растворе заряженных частиц, тем лучше он проводит электрический ток, т.е. чем больше молекул вещества диссоциирует, тем более сильным электролитом оно является.
Список сильных и слабых электролитов нужно знать наизусть!
Сильные электролиты (в растворах): 11
- Растворимые соли
FeCl3, CuSO4, K2CO3 и т. д.
д.
- Щелочи
8 растворимых гидроксидов: LiOH, NaOH, KOH, RbOH, CsOH, Ba(OH)2, Sr(OH)2, Ca(OH)2.
- Сильные кислоты
HI, HBr, HCl, H2SO4(разб), HNO
Слабые электролиты:
- Слабые основания
нерастворимые гидроксиды, NH3∙H2O, растворы аминов
- Слабые кислоты и кислоты средней силы
H3PO4, HF, H2SO3, H2CO3, H2S, H2SiO3, органические кислоты.
- Вода
H2O – очень слабый электролит, диссоциирует ничтожно мало. Чистая дистиллированная вода не проводит ток.
Неэлектролиты: большинство органических соединений, оксиды, вещества, в молекулах которых имеются только ковалентные неполярные или малополярные связи и т.
Сила электролита определяется степенью диссоциации. Рассмотрим соль А2В и кислоту Н3Х:
Диссоциация — всегда обратимый процесс.
Соли диссоциируют (обратимо распадаются на ионы) почти на 100%:
А2В ⇄ 2А+ + В2-. Так как все молекулы распались на ионы, из 1 моль АВ получилось 1 моль В2- и 2 моль А+, то есть три моль ионов.
Многоосновные кислоты и основания диссоциируют ступенчато:
H2X— ⇄ HX2- + H+
HX2-⇄X3- + H+
При этом каждая следующая ступень диссоциации протекает хуже предыдущей, т.к. присутсвует конкурирующий процесс — обратная реакция. Порядок примерно такой: Из 1 моль молекул слабой кислоты по первой ступени диссоциировало 0,05 моль, по второй — 0,0002 моль и по третьей – 0,00000001 моль. Итого образовалось чуть больше 0,1 моль ионов.
Очевидно, этот раствор этой кислоты проводит ток хуже, чем раствор соли.
Пара вопросов для тренировки:
1) Какие частицы образутся при диссоциации нитрата натрия
а) Na+, N+5, O-2; б) Na+, NO3— в) Na, NO2, O2 г) NaNO2, O2
Решение: нитрат натрия образован остатком азотной кислоты и катионом натрия. Уравнение его диссоциации: NaNO3 ⇄ Na+ + NO3—. Ответ б).
2) В четырех пробирках находятся одномолярные растворы следующих веществ:
а) H3PO4 б) Na2SO4 в) NaCl г) HBr
В какой пробирке больше всего ионов?
Решение: a) ортофосфорная кислота – средней силы, диссоциирует слабо, большая часть молекул останутся в растворе молекулами.
б) сульфат натрия – соль, диссоциирует полностью, из одного моль соли олучается три моль ионов: Na2SO4 ⇄ 2Na+ + SO42-.
в) хлорид натрия – соль, диссоциирует полностью, из одного моль соли образуется два моль ионов: NaCl ⇄ Na+ + Cl—.
г) бромоводородная кислота – сильная, но диссоциирует не полностью (в отличие от солей). В реакции HBr ⇄ H+ + Br- из одного моль HBr образуется меньше двух моль ионов.
Ответ б).
Спасибо за то, что пользуйтесь нашими статьями. Информация на странице «Электролитическая диссоциация. Электролиты» подготовлена нашими авторами специально, чтобы помочь вам в освоении предмета и подготовке к экзаменам. Чтобы успешно сдать необходимые и поступить в ВУЗ или техникум нужно использовать все инструменты: учеба, контрольные, олимпиады, онлайн-лекции, видеоуроки, сборники заданий. Также вы можете воспользоваться другими материалами из данного раздела.
Публикация обновлена: 06.07.2023
Киназа контрольной точки внутри-S фазы Chk1 диссоциирует белки репликации Treslin и TopBP1 с помощью нескольких механизмов во время стресса репликации
Сохранить цитату в файл
Формат: Резюме (текст)PubMedPMIDAbstract (текст)CSV
Добавить в коллекции
- Создать новую коллекцию
- Добавить в существующую коллекцию
Имя должно содержать менее 100 символов
Выберите коллекцию: Не удалось загрузить вашу коллекцию из-за ошибки
Повторите попытку
Добавить в мою библиографию
- Моя библиография
Не удалось загрузить делегатов из-за ошибки
Повторите попытку
Ваш сохраненный поиск
Название сохраненного поиска:
Условия поиска:
Тестовые условия поиска
Электронная почта: (изменить)
Который день? Первое воскресеньеПервый понедельникПервый вторникПервая средаПервый четвергПервая пятницаПервая субботаПервый деньПервый рабочий день
Который день? ВоскресеньеПонедельникВторникСредаЧетвергПятницаСуббота
Формат отчета: SummarySummary (text)AbstractAbstract (text)PubMed
Отправить максимум:
1 шт. 5 шт. 10 шт. 20 шт. 50 шт. 100 шт. 200 шт.
5 шт. 10 шт. 20 шт. 50 шт. 100 шт. 200 шт.
Отправить, даже если нет новых результатов
Необязательный текст в электронном письме:
Создайте файл для внешнего программного обеспечения для управления цитированием
. 2022 апр;298(4):101777. doi: 10.1016/j.jbc.2022.101777. Epub 2022 26 февраля.Ребекка Л. Келли 1 , Амелия М Хьюлс 2 , Аннапурна Венкатачалам 1 , Кэтрин Дж. Хантун 2 , Юичи Дж. Мачида 3 , Ларри М. Карниц 4
Принадлежности
- 1 Отделение молекулярной фармакологии и экспериментальной терапии, клиника Майо, Рочестер, Миннесота, США.

- 2 Отдел онкологических исследований, клиника Майо, Рочестер, Миннесота, США.
- 3 Отделение молекулярной фармакологии и экспериментальной терапии, клиника Майо, Рочестер, Миннесота, США; Отдел онкологических исследований, клиника Майо, Рочестер, Миннесота, США.
- 4 Отделение молекулярной фармакологии и экспериментальной терапии, клиника Майо, Рочестер, Миннесота, США; Отдел онкологических исследований, клиника Майо, Рочестер, Миннесота, США. Электронный адрес: [email protected].
- PMID: 35231445
- PMCID: PMC8965152
- DOI:
10.
 1016/j.jbc.2022.101777
1016/j.jbc.2022.101777
Ребекка Л. Келли и др. Дж. Биол. Хим. 2022 Апрель
Бесплатная статья ЧВК . 2022 апрель; 298(4):101777. doi: 10.1016/j.jbc.2022.101777. Epub 2022 26 февраля.Авторы
Ребекка Л. Келли 1 , Амелия М Хьюлс 2 , Аннапурна Венкатачалам 1 , Кэтрин Дж. Хантун 2 , Юичи Дж. Мачида 3 , Ларри М. Карниц 4
Принадлежности
- 1 Отделение молекулярной фармакологии и экспериментальной терапии, клиника Майо, Рочестер, Миннесота, США.

- 2 Отдел онкологических исследований, клиника Майо, Рочестер, Миннесота, США.
- 3 Отделение молекулярной фармакологии и экспериментальной терапии, клиника Майо, Рочестер, Миннесота, США; Отдел онкологических исследований, клиника Майо, Рочестер, Миннесота, США.
- 4 Отделение молекулярной фармакологии и экспериментальной терапии, клиника Майо, Рочестер, Миннесота, США; Отдел онкологических исследований, клиника Майо, Рочестер, Миннесота, США. Электронный адрес: [email protected].
- PMID: 35231445
- PMCID: PMC8965152
- DOI:
10.
 1016/j.jbc.2022.101777
1016/j.jbc.2022.101777
Абстрактный
Стресс репликации препятствует прогрессированию ДНК-полимеразы, вызывая активацию атаксии-телеангиэктазии и связанного с Rad3 сигнального пути, который способствует активности контрольной точки внутри S-фазы посредством фосфорилирования киназы контрольной точки 1 (Chk1). Chk1 подавляет активацию точки начала репликации, частично, путем нарушения взаимодействия между компонентами преинициаторного комплекса Treslin и TopBP1, взаимодействия, которое опосредовано связыванием домена BRCT TopBP1 с двумя сайтами фосфорилирования циклинзависимой киназы (CDK), T968 и S1000 в Треслине. В литературе были предложены две неисключительные модели того, как Chk1 регулирует взаимодействие Treslin-TopBP1: в одной модели эти белки диссоциируют из-за вызванного Chk1 снижения активности CDK, что снижает фосфорилирование сайтов Treslin, связывающих TopBP1, а во второй модели Chk1 непосредственно фосфорилирует Treslin, что приводит к диссоциации TopBP1. Однако формально эти модели не рассматривались. Здесь мы показываем, что фосфорилирование Treslin T968 снижалось зависимым от Chk1 образом, в то время как фосфорилирование Treslin S1000 оставалось неизменным, демонстрируя, что T968 и S1000 регулируются по-разному. Тем не менее, CDK2-опосредованное фосфорилирование само по себе не полностью объясняет регуляцию Chk1 взаимодействия Treslin-TopBP1. Мы также идентифицировали дополнительные сайты фосфорилирования Chk1 на Treslin, которые способствовали нарушению взаимодействия Treslin-TopBP1, включая S1114. Наконец, мы показали, что оба предложенных механизма регулируют активацию ориджина в моделях линий раковых клеток, подвергающихся репликативному стрессу, при этом относительная роль каждого механизма варьируется среди клеточных линий. Это исследование демонстрирует, что Chk1 регулирует Treslin с помощью множества механизмов, способствующих эффективной диссоциации Treslin и TopBP1, и способствует нашему пониманию регуляции Treslin во время контрольной точки внутри S-фазы.
Однако формально эти модели не рассматривались. Здесь мы показываем, что фосфорилирование Treslin T968 снижалось зависимым от Chk1 образом, в то время как фосфорилирование Treslin S1000 оставалось неизменным, демонстрируя, что T968 и S1000 регулируются по-разному. Тем не менее, CDK2-опосредованное фосфорилирование само по себе не полностью объясняет регуляцию Chk1 взаимодействия Treslin-TopBP1. Мы также идентифицировали дополнительные сайты фосфорилирования Chk1 на Treslin, которые способствовали нарушению взаимодействия Treslin-TopBP1, включая S1114. Наконец, мы показали, что оба предложенных механизма регулируют активацию ориджина в моделях линий раковых клеток, подвергающихся репликативному стрессу, при этом относительная роль каждого механизма варьируется среди клеточных линий. Это исследование демонстрирует, что Chk1 регулирует Treslin с помощью множества механизмов, способствующих эффективной диссоциации Treslin и TopBP1, и способствует нашему пониманию регуляции Treslin во время контрольной точки внутри S-фазы.
Ключевые слова: Чк1; ингибиторы Chk1; ТопВР1; Треслин; цитарабин; исходный обжиг; стресс репликации.
Copyright © 2022 Авторы. Опубликовано Elsevier Inc. Все права защищены.
Заявление о конфликте интересов
Конфликт интересов Авторы заявляют об отсутствии конфликта интересов с содержанием данной статьи.
Цифры
Рисунок 1
Chk1 регулирует взаимодействие Treslin–TopBP1…
Рисунок 1
Chk1 регулирует взаимодействие Treslin-TopBP1 . A , клетки K562 были обработаны для…
Рисунок 1 Chk1 регулирует взаимодействие Treslin–TopBP1 .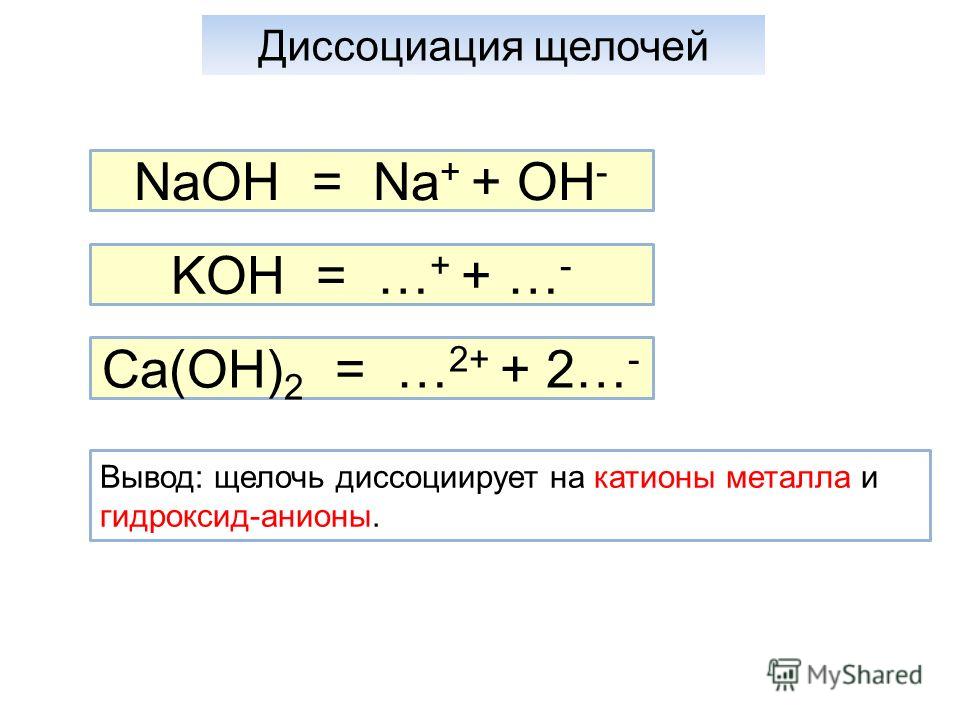 A , клетки K562 перед лизисом обрабатывали в течение 4 ч указанными агентами. Лизаты подвергали иммунопреципитации (IP) с контрольным антителом мышиного иммуноглобулина G (IgG) или моноклональным антителом Treslin (mAb), предварительно связанным с агарозными гранулами белка G. Иммунопреципитаты и лизаты подвергали иммуноблотингу на указанные антигены. B , клетки K562 трансфицировали пустым вектором (EV) или pIRES SFB Treslin WT, инкубировали в течение 24 часов, обрабатывали указанными агентами в течение 4 часов и лизировали. Лизаты инкубировали со стрептавидин-агарозными гранулами для вытягивания SFB Treslin. Пулдауны и лизаты подвергали иммуноблотингу на указанные антигены. C , как в B, но с указанными условиями обработки. * неспецифический диапазон. AraC, цитарабин; Chk1, киназа 1 контрольной точки; SFB, S-Tag, FLAG, стрептавидин-связывающий пептид.
A , клетки K562 перед лизисом обрабатывали в течение 4 ч указанными агентами. Лизаты подвергали иммунопреципитации (IP) с контрольным антителом мышиного иммуноглобулина G (IgG) или моноклональным антителом Treslin (mAb), предварительно связанным с агарозными гранулами белка G. Иммунопреципитаты и лизаты подвергали иммуноблотингу на указанные антигены. B , клетки K562 трансфицировали пустым вектором (EV) или pIRES SFB Treslin WT, инкубировали в течение 24 часов, обрабатывали указанными агентами в течение 4 часов и лизировали. Лизаты инкубировали со стрептавидин-агарозными гранулами для вытягивания SFB Treslin. Пулдауны и лизаты подвергали иммуноблотингу на указанные антигены. C , как в B, но с указанными условиями обработки. * неспецифический диапазон. AraC, цитарабин; Chk1, киназа 1 контрольной точки; SFB, S-Tag, FLAG, стрептавидин-связывающий пептид.
Рисунок 2
Активация Chk1 уменьшает Treslin T968…
Рисунок 2
Активация Chk1 снижает фосфорилирование Treslin T968, но не S1000 . А , схема…
А , схема…
Активация Chk1 снижает фосфорилирование Treslin T968, но не S1000 . A , диаграмма SFB Treslin (полноразмерный Treslin, аминокислоты 1–1910), SFB Treslin Δ900–1100 (отсутствуют аминокислоты 900–1100) и SFB Treslin 900–1100 (только аминокислоты 900–1100). Относительное положение последовательности RNL, сайтов T968 и S1000 и области связывания TopBP1 ( пунктирный прямоугольник ) указаны на полноразмерном белке. B , клетки K562 трансфицировали WT pIRES SFB Treslin 900–1100 и обозначали мутанты, инкубировали в течение 24 часов и лизировали. Лизаты инкубировали со стрептавидин-агарозными гранулами для извлечения фрагментов SFB Treslin. Пулдауны и лизаты подвергали иммуноблотингу на указанные антигены. C , клетки К562 трансфицировали pIRES SFB Treslin 900–1100 и pIRES SFB Treslin Δ900–1100, инкубировали в течение 24 ч, обрабатывали указанными агентами в течение 4 ч и лизировали. Лизаты инкубировали со стрептавидин-агарозными гранулами для вытягивания фрагментов Треслина SFB. Пулдауны и лизаты подвергали иммуноблотингу на указанные антигены, включая фосфо-Т9.68 Треслин (pTreslin T968) и фосфо-S1000 Treslin (pTreslin S1000). D , клетки К562 перед лизисом обрабатывали в течение 4 ч указанными агентами. Лизаты инкубировали с гранулами агарозы с белком G, предварительно связанными с контрольным мышиным моноклональным антителом IgG или моноклональным антителом Treslin (mAb). Иммунопреципитаты и лизаты подвергали иммуноблотингу на указанные антигены. AraC, цитарабин; Chk1, киназа 1 контрольной точки; SFB, S-Tag, FLAG, стрептавидин-связывающий пептид; РНЛ, аргинин-аспарагин-лейцин.
Лизаты инкубировали со стрептавидин-агарозными гранулами для вытягивания фрагментов Треслина SFB. Пулдауны и лизаты подвергали иммуноблотингу на указанные антигены, включая фосфо-Т9.68 Треслин (pTreslin T968) и фосфо-S1000 Treslin (pTreslin S1000). D , клетки К562 перед лизисом обрабатывали в течение 4 ч указанными агентами. Лизаты инкубировали с гранулами агарозы с белком G, предварительно связанными с контрольным мышиным моноклональным антителом IgG или моноклональным антителом Treslin (mAb). Иммунопреципитаты и лизаты подвергали иммуноблотингу на указанные антигены. AraC, цитарабин; Chk1, киназа 1 контрольной точки; SFB, S-Tag, FLAG, стрептавидин-связывающий пептид; РНЛ, аргинин-аспарагин-лейцин.
Рисунок 3
Сохранение фосфорилирования Treslin T968…
Рисунок 3
Сохранение фосфорилирования Treslin T968 не предотвращает диссоциацию Treslin-TopBP1 .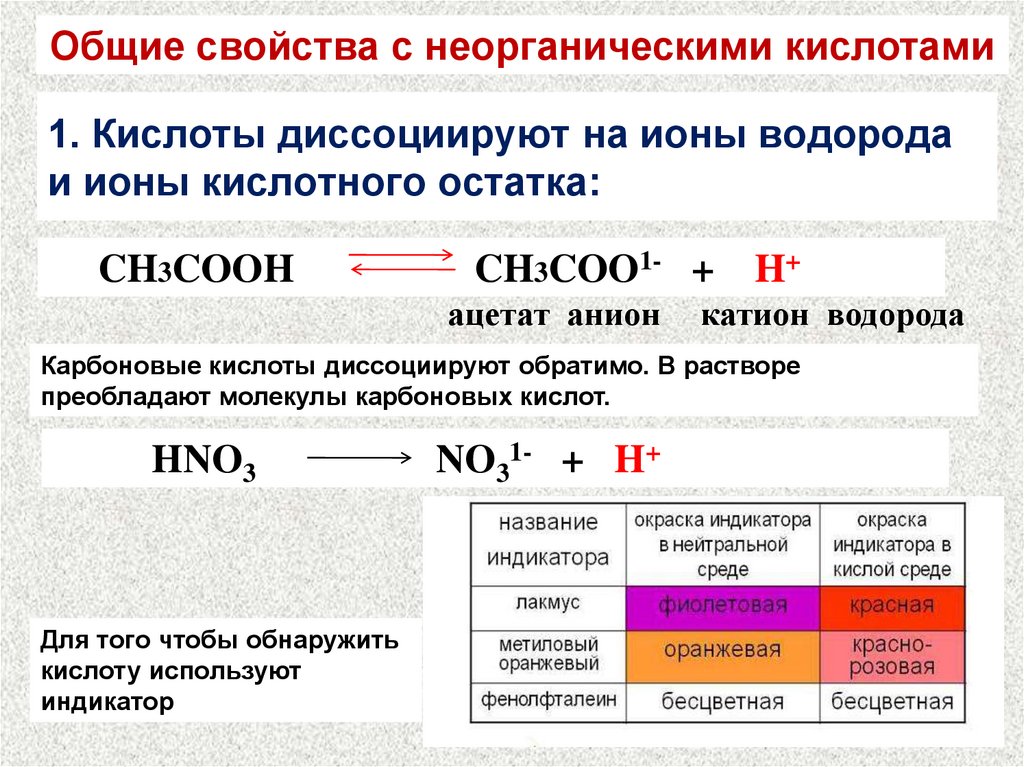 А ,…
А ,…
Сохранение Treslin T968 фосфорилирование не предотвращает диссоциацию Treslin-TopBP1 . A , клетки K562 котрансфицировали пустым вектором (EV), pIRES SFB Treslin WT или pIRES SFB Treslin 900–1100 с pCMV CDK2 HA AF или без него (как указано), инкубировали в течение 24 ч, обрабатывали 10 мкМ AraC в течение 4 часов и лизируется. Лизаты инкубировали со стрептавидин-агарозными гранулами для удаления белков, меченных SFB. Лизаты и белки, связанные с шариками, подвергали иммуноблотингу на указанные антигены. B , клетки K562 трансфицировали пустым вектором (EV), pCMV CDK2 HA WT или pCMV CDK2 HA AF, инкубировали в течение 24 часов, обрабатывали 10 мкМ AraC в течение 4 часов и лизировали. Лизаты инкубировали с агарозными шариками с белком G, предварительно связанными с контрольным антителом IgG мыши или моноклональным антителом Treslin (mAb). Лизаты и белки, связанные с шариками, подвергали иммуноблотингу на указанные антигены. C , как в B, но клетки K562 трансфицировали пустым вектором (EV), pIRES SFB Treslin WT или pIRES SFB Treslin 7A вместе с pCMV CDK2 HA WT или pCMV CDK2 HA AF, как указано. CDK, циклинзависимая киназа; AraC, цитарабин; Chk1, киназа 1 контрольной точки; SFB, S-Tag, FLAG, стрептавидин-связывающий пептид.
C , как в B, но клетки K562 трансфицировали пустым вектором (EV), pIRES SFB Treslin WT или pIRES SFB Treslin 7A вместе с pCMV CDK2 HA WT или pCMV CDK2 HA AF, как указано. CDK, циклинзависимая киназа; AraC, цитарабин; Chk1, киназа 1 контрольной точки; SFB, S-Tag, FLAG, стрептавидин-связывающий пептид.
Рисунок 4
С-клемма Treslin Chk1…
Рисунок 4
С-концевой связывающий домен Treslin Chk1 регулирует взаимодействие Treslin-TopBP1, способствуя фосфорилированию…
Рисунок 4 С-концевой связывающий домен Treslin Chk1 регулирует взаимодействие Treslin-TopBP1, способствуя фосфорилированию Treslin вблизи области связывания TopBP1 . A , схема, изображающая белок Treslin, сайты фосфорилирования T968 и S1000 ( зеленый ), консенсусные сайты фосфорилирования Chk1 ( красный ; S937, S1019, S1025, S1038, S1044 и S1114) и С-концевая область взаимодействия Chk1 ( Chk1 ). B , клетки K562 перед лизисом обрабатывали в течение 4 ч указанными агентами. Лизаты инкубировали с гранулами агарозы с белком G, предварительно связанными с моноклональным антителом Treslin (mAb) для иммунопреципитации (IP). Связанные с шариками белки подвергали иммуноблотингу на фосфо-S1114 Treslin (pTreslin S1114) и Treslin. C, клетки K562 трансфицировали пустым вектором (EV), pIRES SFB Treslin WT или pIRES SFB Treslin 7A, инкубировали в течение 24 часов, обрабатывали 10 мкМ AraC в течение 4 часов и лизировали. Лизаты инкубировали со стрептавидин-агарозой для удаления белков, меченных SFB. Лизаты и белки, связанные с шариками, подвергали иммуноблотингу на указанные антигены. D , клетки K562 котрансфицировали либо пустым вектором, pIRES SFB Treslin WT, либо pIRES SFB Treslin S6A вместе с pCMV CDK2 HA WT или pCMV CDK2 HA AF, как указано, инкубировали в течение 24 часов, обрабатывали в течение 4 часов с 10 мкМ AraC и лизировали. Лизаты инкубировали со стрептавидин-агарозными гранулами для удаления белков, меченных SFB.
B , клетки K562 перед лизисом обрабатывали в течение 4 ч указанными агентами. Лизаты инкубировали с гранулами агарозы с белком G, предварительно связанными с моноклональным антителом Treslin (mAb) для иммунопреципитации (IP). Связанные с шариками белки подвергали иммуноблотингу на фосфо-S1114 Treslin (pTreslin S1114) и Treslin. C, клетки K562 трансфицировали пустым вектором (EV), pIRES SFB Treslin WT или pIRES SFB Treslin 7A, инкубировали в течение 24 часов, обрабатывали 10 мкМ AraC в течение 4 часов и лизировали. Лизаты инкубировали со стрептавидин-агарозой для удаления белков, меченных SFB. Лизаты и белки, связанные с шариками, подвергали иммуноблотингу на указанные антигены. D , клетки K562 котрансфицировали либо пустым вектором, pIRES SFB Treslin WT, либо pIRES SFB Treslin S6A вместе с pCMV CDK2 HA WT или pCMV CDK2 HA AF, как указано, инкубировали в течение 24 часов, обрабатывали в течение 4 часов с 10 мкМ AraC и лизировали. Лизаты инкубировали со стрептавидин-агарозными гранулами для удаления белков, меченных SFB. Лизаты и белки, связанные с шариками, подвергали иммуноблотингу на указанные антигены. * неспецифический диапазон. E . Левая панель: во время возбуждения ориджина Treslin и TopBP1 взаимодействуют через N-концевые домены BRCT TopBP1, которые связывают два сайта фосфорилирования на Treslin (T968 и S1000). Правая панель: Репликационный стресс активирует Chk1, который имеет два механизма, влияющих на взаимодействие Treslin-TopBP1: № 1 — Chk1 ингибирует активность CDK посредством истощения и/или секвестрации фосфатаз семейства CDC25, № 2 — Chk1 связывается непосредственно с С-концом Treslin. и фосфорилирует сайты Chk1 на Treslin. Ингибитор Chk1 блокирует оба механизма. AraC, цитарабин; Chk1, киназа 1 контрольной точки; SFB, S-Tag, FLAG, стрептавидин-связывающий пептид; CDK, циклинзависимая киназа.
Лизаты и белки, связанные с шариками, подвергали иммуноблотингу на указанные антигены. * неспецифический диапазон. E . Левая панель: во время возбуждения ориджина Treslin и TopBP1 взаимодействуют через N-концевые домены BRCT TopBP1, которые связывают два сайта фосфорилирования на Treslin (T968 и S1000). Правая панель: Репликационный стресс активирует Chk1, который имеет два механизма, влияющих на взаимодействие Treslin-TopBP1: № 1 — Chk1 ингибирует активность CDK посредством истощения и/или секвестрации фосфатаз семейства CDC25, № 2 — Chk1 связывается непосредственно с С-концом Treslin. и фосфорилирует сайты Chk1 на Treslin. Ингибитор Chk1 блокирует оба механизма. AraC, цитарабин; Chk1, киназа 1 контрольной точки; SFB, S-Tag, FLAG, стрептавидин-связывающий пептид; CDK, циклинзависимая киназа.
Рисунок 5
Комбинированное выражение Treslin S6A…
Рисунок 5
Комбинированная экспрессия Treslin S6A и CDK2 AF способствует синтезу и происхождению ДНК…
Рисунок 5 Комбинированная экспрессия Treslin S6A и CDK2 AF способствует синтезу ДНК и запуску ориджина в обработанных AraC клетках K562. A , схема анализа включения EdU. Клетки обрабатывали AraC в течение 3 ч с добавлением EdU в течение последнего часа обработки. B , графики рассеяния из эксперимента по включению EdU. Клетки K562 трансфицировали пустым вектором (EV) и указанными плазмидами, культивировали в течение 24 часов и не обрабатывали ничем (NT) или 0,1 мкМ AraC в течение 3 часов. 10 мкМ EdU добавляли в течение последнего часа обработки. Затем образцы собирали, фиксировали и окрашивали Alexafluor 594 для EdU и FxCycle Violet для содержания ДНК. Gates указывают на EdU-положительную клеточную популяцию. C , гистограмма показывает процент EdU-позитивных клеток в условиях обработки AraC, показанных в B . Значения представляют собой среднее значение трех независимых экспериментов со стандартной ошибкой (среднее значение ± стандартная ошибка среднего). Односторонний ANOVA с тестом множественных сравнений Тьюки был выполнен для оценки влияния экспрессии SFB Treslin и CDK2 HA на включение EdU после обработки AraC (∗∗∗ p -значение <0,0005).
A , схема анализа включения EdU. Клетки обрабатывали AraC в течение 3 ч с добавлением EdU в течение последнего часа обработки. B , графики рассеяния из эксперимента по включению EdU. Клетки K562 трансфицировали пустым вектором (EV) и указанными плазмидами, культивировали в течение 24 часов и не обрабатывали ничем (NT) или 0,1 мкМ AraC в течение 3 часов. 10 мкМ EdU добавляли в течение последнего часа обработки. Затем образцы собирали, фиксировали и окрашивали Alexafluor 594 для EdU и FxCycle Violet для содержания ДНК. Gates указывают на EdU-положительную клеточную популяцию. C , гистограмма показывает процент EdU-позитивных клеток в условиях обработки AraC, показанных в B . Значения представляют собой среднее значение трех независимых экспериментов со стандартной ошибкой (среднее значение ± стандартная ошибка среднего). Односторонний ANOVA с тестом множественных сравнений Тьюки был выполнен для оценки влияния экспрессии SFB Treslin и CDK2 HA на включение EdU после обработки AraC (∗∗∗ p -значение <0,0005). D , схема мечения волокон ДНК. Клетки К562 трансфицировали указанными плазмидами, культивировали в течение 24 ч, импульсно метили IdU ( красный ), промыли, обработали ничем или 0,5 мкМ афидиколином (APH) и обработали CldU ( зеленый ). E показаны репрезентативные изображения для WT/WT, NT, WT/WT +APH и S6A/AF, +APH. Показана белая стрелка , указывающая на новое происхождение. F гистограмма показывает процентное соотношение новых источников по сравнению с общими событиями, подсчитанными в каждом состоянии в E. Для каждого состояния было подсчитано не менее 300 событий. Значения представляют собой среднее значение трех независимых экспериментов со стандартной ошибкой (среднее значение ± стандартная ошибка среднего). Двухфакторный дисперсионный анализ с тестом множественных сравнений Тьюки был проведен для оценки влияния экспрессии SFB Treslin и CDK2 HA на возбуждение происхождения. (∗∗∗∗ p -значение < 0,0001). AraC, цитарабин; Chk1, киназа 1 контрольной точки; SFB, S-Tag, FLAG, стрептавидин-связывающий пептид; CDK, циклинзависимая киназа.
D , схема мечения волокон ДНК. Клетки К562 трансфицировали указанными плазмидами, культивировали в течение 24 ч, импульсно метили IdU ( красный ), промыли, обработали ничем или 0,5 мкМ афидиколином (APH) и обработали CldU ( зеленый ). E показаны репрезентативные изображения для WT/WT, NT, WT/WT +APH и S6A/AF, +APH. Показана белая стрелка , указывающая на новое происхождение. F гистограмма показывает процентное соотношение новых источников по сравнению с общими событиями, подсчитанными в каждом состоянии в E. Для каждого состояния было подсчитано не менее 300 событий. Значения представляют собой среднее значение трех независимых экспериментов со стандартной ошибкой (среднее значение ± стандартная ошибка среднего). Двухфакторный дисперсионный анализ с тестом множественных сравнений Тьюки был проведен для оценки влияния экспрессии SFB Treslin и CDK2 HA на возбуждение происхождения. (∗∗∗∗ p -значение < 0,0001). AraC, цитарабин; Chk1, киназа 1 контрольной точки; SFB, S-Tag, FLAG, стрептавидин-связывающий пептид; CDK, циклинзависимая киназа.
Рисунок 6
Только Treslin S6A способствует продвижению ДНК…
Рисунок 6
Треслин S6A сам по себе способствует синтезу ДНК и начальному возбуждению в клетках U2OS. А…
Рисунок 6 Treslin S6A сам по себе способствует синтезу ДНК и активизации происхождения в клетках U2OS. A , клетки U2OS трансфицировали пустым вектором (EV), SFB-меченым Treslin дикого типа (WT) или Treslin S6A (S6A), культивировали в течение 48 ч, не обрабатывали ничем (-) или 10 мкМ AraC в течение 2 ч , и лизируется. Клеточные лизаты инкубировали со стрептавидин-агарозными гранулами для вытягивания белков, меченных SFB. Пуллдауны проводили на геле и проводили иммуноблотинг для указанных антигенов. Все образцы были обработаны на одном и том же геле, а посторонние дорожки были вырезаны из изображений иммуноблотов и обозначены штрихами. B , клеточные линии TRE3G U2OS, трансдуцированные меченым HA Treslin дикого типа (WT) или Treslin S6A (S6A), меченным HA, обрабатывали в течение 24 ч без (-) или 10 нг/мл (+) доксициклина (Dox) для индуцировать экспрессию HA Treslin. Лизаты клеток подвергали иммуноблотингу на указанные антигены. * неспецифический диапазон. C , анализ включения EdU, как показано на схематической фигуре. 5 Анализ проводили с клеточными линиями TRE3G U2OS. Клетки U2OS обрабатывали 10 нг/мл доксициклина, культивировали в течение 24 часов и не обрабатывали ничем (без обработки, NT) или 0,1 мкМ AraC в течение 3 часов. 10 мкМ EdU добавляли в течение последнего часа обработки. Затем образцы собирали, фиксировали и окрашивали для включения EdU (Alexafluor 59).4) и содержание ДНК (FxCycle Violet). D , гистограмма показывает процент EdU-положительных клеток в каждом состоянии из C. Значения представляют собой среднее значение трех независимых экспериментов со стандартной ошибкой (среднее ± SEM).
B , клеточные линии TRE3G U2OS, трансдуцированные меченым HA Treslin дикого типа (WT) или Treslin S6A (S6A), меченным HA, обрабатывали в течение 24 ч без (-) или 10 нг/мл (+) доксициклина (Dox) для индуцировать экспрессию HA Treslin. Лизаты клеток подвергали иммуноблотингу на указанные антигены. * неспецифический диапазон. C , анализ включения EdU, как показано на схематической фигуре. 5 Анализ проводили с клеточными линиями TRE3G U2OS. Клетки U2OS обрабатывали 10 нг/мл доксициклина, культивировали в течение 24 часов и не обрабатывали ничем (без обработки, NT) или 0,1 мкМ AraC в течение 3 часов. 10 мкМ EdU добавляли в течение последнего часа обработки. Затем образцы собирали, фиксировали и окрашивали для включения EdU (Alexafluor 59).4) и содержание ДНК (FxCycle Violet). D , гистограмма показывает процент EdU-положительных клеток в каждом состоянии из C. Значения представляют собой среднее значение трех независимых экспериментов со стандартной ошибкой (среднее ± SEM). Односторонний ANOVA с тестом множественных сравнений Тьюки был выполнен для оценки влияния HA Treslin на включение EdU после обработки AraC (∗∗∗∗ p -значение <0,0001). E , маркировку волокон ДНК проводили, как показано на схеме на рис. 5 D . Клетки U2OS обрабатывали 10 нг/мл доксициклина за 24 часа до анализа. Клетки метили в импульсном режиме с помощью IdU (, красный ), промывали, не обрабатывали ничем или 5 мкМ афидиколином (APH) и импульсно метили CldU (, зеленый ). Показаны репрезентативные изображения данных. Белые стрелки указывают на новое происхождение. F , столбчатая диаграмма показывает процент нового происхождения в каждом состоянии из E. Значения представляют собой среднее значение трех независимых экспериментов со стандартной ошибкой (среднее ± SEM). Двухфакторный дисперсионный анализ с тестом множественных сравнений Тьюки был проведен для оценки влияния экспрессии HA Treslin на возбуждение происхождения. (∗∗ p -значение < 0,005).
Односторонний ANOVA с тестом множественных сравнений Тьюки был выполнен для оценки влияния HA Treslin на включение EdU после обработки AraC (∗∗∗∗ p -значение <0,0001). E , маркировку волокон ДНК проводили, как показано на схеме на рис. 5 D . Клетки U2OS обрабатывали 10 нг/мл доксициклина за 24 часа до анализа. Клетки метили в импульсном режиме с помощью IdU (, красный ), промывали, не обрабатывали ничем или 5 мкМ афидиколином (APH) и импульсно метили CldU (, зеленый ). Показаны репрезентативные изображения данных. Белые стрелки указывают на новое происхождение. F , столбчатая диаграмма показывает процент нового происхождения в каждом состоянии из E. Значения представляют собой среднее значение трех независимых экспериментов со стандартной ошибкой (среднее ± SEM). Двухфакторный дисперсионный анализ с тестом множественных сравнений Тьюки был проведен для оценки влияния экспрессии HA Treslin на возбуждение происхождения. (∗∗ p -значение < 0,005).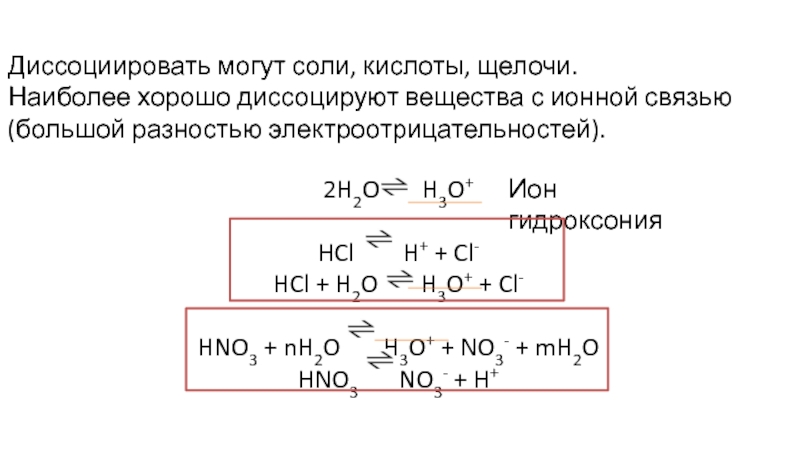 AraC, цитарабин; Chk1, киназа 1 контрольной точки; SFB, S-Tag, FLAG, стрептавидин-связывающий пептид.
AraC, цитарабин; Chk1, киназа 1 контрольной точки; SFB, S-Tag, FLAG, стрептавидин-связывающий пептид.
См. это изображение и информацию об авторских правах в PMC
Похожие статьи
- Непосредственная роль репликационного белка треслина (Ticrr) в ответе контрольной точки, опосредованном киназой ATR.
Хассан Б.Х., Линдси-Больц Л.А., Кемп М.Г., Санкар А. Хассан Б.Х. и др. Дж. Биол. Хим. 2013 28 июня; 288(26):18903-10. doi: 10.1074/jbc.M113.475517. Epub 2013 21 мая. Дж. Биол. Хим. 2013. PMID: 23696651 Бесплатная статья ЧВК.
- Мутантный p53 нарушает контроль контрольной точки репликации ДНК через TopBP1 и Treslin.
Liu K, Lin FT, Graves JD, Lee YJ, Lin WC. Лю К. и др. Proc Natl Acad Sci U S A. 9 мая 2017 г .
 ; 114 (19): E3766-E3775. doi: 10.1073/pnas.1619832114. Epub 2017 24 апр.
Proc Natl Acad Sci U S A. 2017.
PMID: 28439015
Бесплатная статья ЧВК.
; 114 (19): E3766-E3775. doi: 10.1073/pnas.1619832114. Epub 2017 24 апр.
Proc Natl Acad Sci U S A. 2017.
PMID: 28439015
Бесплатная статья ЧВК. - Прямая регуляция Treslin циклинзависимой киназой необходима для начала репликации ДНК.
Кумагаи А., Шевченко А., Шевченко А., Данфи В.Г. Кумагаи А. и др. Джей Селл Биол. 2011 13 июня; 193 (6): 995-1007. doi: 10.1083/jcb.201102003. Epub 2011 6 июня. Джей Селл Биол. 2011. PMID: 21646402 Бесплатная статья ЧВК.
- TopBP1 и альфа-опосредованное ДНК-полимеразой привлечение комплекса 9-1-1 к остановленным вилкам репликации: последствия для механизма, основанного на перезапуске репликации, для активации контрольной точки ATR.
Ян С., Майкл В.М. Ян С. и др. Клеточный цикл. 2009 15 сентября; 8 (18): 2877-84.
 doi: 10.4161/cc.8.18.9485. Epub 2009 9 сентября.
Клеточный цикл. 2009.
PMID: 19652550
Обзор.
doi: 10.4161/cc.8.18.9485. Epub 2009 9 сентября.
Клеточный цикл. 2009.
PMID: 19652550
Обзор. - Передача сигналов спящего происхождения во время невозмущенной репликации.
Моисеева Т.Н., Баккенист С.Я. Моисеева ТН и соавт. Восстановление ДНК (Amst). 2019 сен;81:102655. doi: 10.1016/j.dnarep.2019.102655. Epub 2019 8 июля. Восстановление ДНК (Amst). 2019. PMID: 31311769 Бесплатная статья ЧВК. Обзор.
Посмотреть все похожие статьи
Рекомендации
- Боос Д., Феррейра П. Правила обжига происхождения для контроля времени репликации генома. Гены (Базель) 2019;10:199. — ЧВК — пабмед
- Миядзава-Онами М.
 , Араки Х., Танака С. Сборка комплекса перед инициацией функционирует как молекулярный переключатель, который расщепляет двойной гексамер Mcm2-7. EMBO Rep. 2017; 18: 1752–1761.
—
ЧВК
—
пабмед
, Араки Х., Танака С. Сборка комплекса перед инициацией функционирует как молекулярный переключатель, который расщепляет двойной гексамер Mcm2-7. EMBO Rep. 2017; 18: 1752–1761.
—
ЧВК
—
пабмед
- Миядзава-Онами М.
- Блейхерт Ф., Ботчан М. Р., Бергер Дж. М. Механизмы инициации репликации клеточной ДНК. Наука. 2017;355 — пабмед
- Кумагаи А.
 , Шевченко А., Шевченко А., Данфи В. Г. Прямая регуляция Treslin циклинзависимой киназой необходима для начала репликации ДНК. Дж. Клеточная биология. 2011;193: 995–1007.
—
ЧВК
—
пабмед
, Шевченко А., Шевченко А., Данфи В. Г. Прямая регуляция Treslin циклинзависимой киназой необходима для начала репликации ДНК. Дж. Клеточная биология. 2011;193: 995–1007.
—
ЧВК
—
пабмед
- Кумагаи А.
- Боос Д., Санчес-Пулидо Л., Рапас М., Перл Л.Х., Оливер А.В., Понтинг С.П., Диффли Дж.Ф. Регуляция репликации ДНК посредством взаимодействия Sld3-Dpb11 сохраняется от дрожжей до человека. Курс. биол. 2011;21:1152–1157. — пабмед
термины MeSH
вещества
Грантовая поддержка
- P30 CA015083/CA/NCI NIH HHS/США
- R01 CA1/CA/NCI NIH HHS/США
- T32 GM072474/GM/NIGMS NIH HHS/США
Укажите
Формат: ААД АПА МДА НЛМ
Отправить наВсе диссоциируют.
 Набор графики Janae Elisabeth… | by Trauma Geek | Эпоха Сознания Все диссоциируют. Набор графики Janae Elisabeth… | by Trauma Geek | Эпоха осознания | Средний
Набор графики Janae Elisabeth… | by Trauma Geek | Эпоха Сознания Все диссоциируют. Набор графики Janae Elisabeth… | by Trauma Geek | Эпоха осознания | СреднийНабор рисунков Яны Элизабет, основанный на поливагальной теории
Опубликовано в·
Чтение через 2 минуты качественная PDF-версия этих рисунков доступна для печати и образовательных целях по телефону TraumaGeek.com .Посмотреть другую инфографику: linktr.ee/TraumaGeek
Профессиональные покровители получают всю коллекцию графики Trauma Geek: www.Patreon.com/TraumaGeek
Диссоциация
Поливагальная теория
Травма
.2K Followers
·Писатель дляДжена Элизабет — исследователь, рассказчик и защитник нейроразнообразия. из западной части Северной Каролины. #actuallyautistic #traumaismyspecialinterest
Еще из Trauma Geek and Age of Awareness
Trauma Geek
Трудности перевода: теория нейродивергенции социального языка (часть 1 из 2)
от Janae Elisabeth
Чтение: 12 мин·23 мая 2020 г. Как мы думаем
Как мы думаем
За последние 50 с лишним лет новый способ мышления становится все более популярным. Сосредоточено на основных человеческих способностях — эмпатии…
9 мин чтения · 22 февраля 2021 г.
Zohvib
in
Я вел дневник каждый день в течение 90 дней. Вот что я узнал
Документирование мелких подробностей вашей повседневной жизни становится праздником того, кто вы есть.
Чтение за 4 минуты · 2 июня
Trauma Geek
Нейродивергентный обзор протокола безопасного и звукового воздействия
Протокол безопасного и звукового воздействия — это мощное вмешательство при травмах, разработанное доктором Стивеном Поргесом и его командой. Доктор Поргес наиболее известен благодаря…
10 минут чтения·9 февраля
Просмотреть все от Trauma Geek
Рекомендовано на Medium
Непристойный
10 секунд, которые закончились моим 20-летним браком
В Северной Вирджинии август, жарко и влажно.
 Я до сих пор не принял душ после утренней пробежки. Я ношу свою домохозяйку… ·Чтение на 4 мин·16 февраля 2022 г.
Я до сих пор не принял душ после утренней пробежки. Я ношу свою домохозяйку… ·Чтение на 4 мин·16 февраля 2022 г.Aleid ter Weel
in
10 вещей, которые можно сделать вечером вместо просмотра Netflix
9000 5Привычки без устройств для повышения продуктивности и счастья.
·5 минут чтения·15 февраля 2022 г.Списки
Лучшее из писательского кооператива
67 рассказов·8 сохранений
Непристойный
How My Hus Сексуальность группы определила наш брак
И ложь, которую я сказал себе
·5 минут чтения·17 маяPyCoach
в
Вы используете ChatGPT неправильно! Вот как опередить 99% пользователей ChatGPT
·7 мин чтения·17 мартаНик Виньялл
3 привычки, которые сделают вас психически сильным
#2: избегайте ненужных мысленных путешествий во времени
9001 4 · 9 минут чтения · 2 апреляZulie Rane
in
Если вы хотите стать творцом, удалите все (кроме двух) платформы социальных сетей
В октябре 2022 года, во время разгрома Илона Маска, я наконец-то удалил Twitter со своего телефона.

