13 Основные теории памяти.
Ассоциативная теория.
Ассоциация — это связь между отдельными представлениями, при которых одно из этих представлений вызывает другое.
Ассоциации образуются на случайной
основе, поэтому ассоциативная теория
не объясняет избирательности памяти.
Тем не менее, ассоциативная теория дала
много полезного для понимания законов
памяти. В рамках этой теории работал Г.
Эббингауз, которому принадлежит открытие
ряда механизмов и закономерностей
памяти. Процесс возрождения некоторого
психического содержания, воспринятого
ранее в виде представлений, Эббингауз
назвал воспроизведением. Механизм
воспроизведения он назвал ассоциацией
— мысленной связью, возникающей между
процессом, наблюдаемым в действительности,
и возможностью его наступления в случае
его отсутствия. Таким образом, ассоциация
— это внутренняя причина воспроизведения.
При этом Эббингауз подчеркивал, что
воспроизводимые ощущения и представления
не тождественны с существовавшими
ранее, а только сходны с ними, и, тем не
менее, способны пробуждать наблюдаемые
ранее душевные образования.
1. по сходству;
2. по контрасту;
3. по временной связи;
4. по пространственному сосуществованию.
Гештальттеория.
Гештальттеория пришла на смену
ассоциативной теории. Исследования
памяти были проведены К.Левином. По
мнению ее создателей, память определяется
законами формирования гештальта
(целостной структуры). В этой связи для
эффективного функционирования памяти
чрезвычайно важным признается
структурирование материала, его
систематизация при запоминании и
воспроизведении. Процессуально память
представлялась так: у человека имеется
некоторая необходимость (потребность),
дающая установку на запоминание или
воспроизведение.
Б.В.Зейгарник она показала, что незавершенные действия приводятся лучше, чем завершенные. Это получше название «эффект зейгарник», или эффект незаконченного действия. Они запоминаются лучше потому, что они связаны с искусствено создаваемой потребностью завершить действие, которое не полунию разрядки, это наз.квазипотребность.
Теория бихевиоризма.
Есть вид поведения, кот.формируется
в процессе многократного повторения
связей между стимулами и реакциями.
Осн.Уотсон, Э.Торондайк, Скиннер. Вместо
слова память часто испол.научение.Память
рассматривалась бихевиористами как
процесс, обеспечивающий научение.
Результативность научения зависела от
качества памяти. Продуктивность же
последней определялась подкреплениями
— как положительными, так и отрицательными.
Таким образом, теория памяти в таком
представлении формировалась как теория
подкреплений.
Когнитивная теория памяти.
Когнитивная психология строится на положении о решающей роли знания, когнитивных структур в протекании психических процессов и поведении человека. Исследуются вопросы организации знания в памяти человека, соотношение вербальных и образных компонентов в процессах запоминания.
Основная идея когнитивной психологии
состоит в неразрывном единстве всех
познавательных психических процессов,
представляющих единую когнитивную
структуру.
Модель Во и Норманна: делит память на первичную (систему кратковременного хранения) и вторичную (систему более длительного хранения). Эта модель спровоцировала представление о памяти как некоем «ящике в голове». Система кратковременного хранения имеет очень ограниченный объем, и информация в ней теряется путем вытеснения новой.
Модель Аткинсона и Шифрина: они ввели в кратковременную и долговременную память гораздо больше подсистем. Чем предыдущие авторы. Их модель была дополнена сенсорным регистром, кратковременным и долговременным хранилищами. При этом дифференцировались понятия «память» (как данные, подлежащие сохранению) и «хранилище» (как структурный элемент, для хранения этих данных).
Психоаналитическая теория.
Важной ее предпосылкой является
зависимость памяти от мотивации и
эмоциональных состояний человека. Согласно Фрейду, из памяти вытесняется
все, что не соответствует подсознательным
влечениям человека, а все приятное
сохраняется. Но эта зависимость не
получила экспериментального подтверждения.
Так, в опытах П.П.Блонского, напротив,
было отмечено, что подавляющее большинство
испытуемых вспоминали события, имеющие
отрицательную эмоциональную окраску.
Согласно Фрейду, из памяти вытесняется
все, что не соответствует подсознательным
влечениям человека, а все приятное
сохраняется. Но эта зависимость не
получила экспериментального подтверждения.
Так, в опытах П.П.Блонского, напротив,
было отмечено, что подавляющее большинство
испытуемых вспоминали события, имеющие
отрицательную эмоциональную окраску.
Биохимическая (молекулярная) теория памяти.
Главный вывод данной теории состоит
в том, что существует материальный
носитель памяти. Таковыми являются
молекулы нуклеиновых кислот РНК и ДНК.
Причем изменения в ДНК связывают с
долговременной памятью, а в РНК — с
оперативной памятью. Эти данные были
получены опытным путем. Содержание
опытов заключалось в следующем: у крыс
вырабатывался определенный условный
рефлекс (-заданное поведение в Т-образном
лабиринте), после чего из их мозга было
выделено белковое вещество и внедрено
в мозг необученных крыс, которые
подверглись аналогичному обучению.
Выработка соответствующего рефлекса
у необученных крыс происходила заметно
быстрее, чем у предшествующей группы.
Нейронная теория памяти.
Нейронная теория основывается на
традициях школы физиолога И.П.Павлова.
Согласно ей, в основе памяти лежит
свойство нервной ткани изменяться под
влиянием действия раздражителей и
сохранять в себе следы нервного
возбуждения. Под следами в данном случае
понимают определенные электрические
и механические изменения в синапсах.
Эти следы могут при определенных условиях
актуализироваться (оживляться). При
отсутствии раздражителя, вызвавшего
указанные изменения, в них возникает
процесс возбуждения, который и
обусловливает возникновение представлений
(образов памяти).
Деятельностная теория памяти.
В контексте этой теории память
выступает как особая психическая
деятельность, направленная на решение
определенной мнемической задачи. Память
как деятельность включает действия
запоминания, сохранения и воспроизведения.
Будучи деятельностью память имеет все
ее структурные элементы, в частности —
цель, средства, операции. А результат
мнемической деятельности всегда зависит
от ее организации (П.И.Зинченко,
А.Н.Леонтьев, А.А.Смирнов). П.И.Зинченко
показал, что слова, кодируемые значащими
средствами, будут сохраняться лучше,
чем слова, кодируемые более поверхностными
средствами. Запоминаемость слов сильно
зависит от цели, стоящей при усвоении
материала.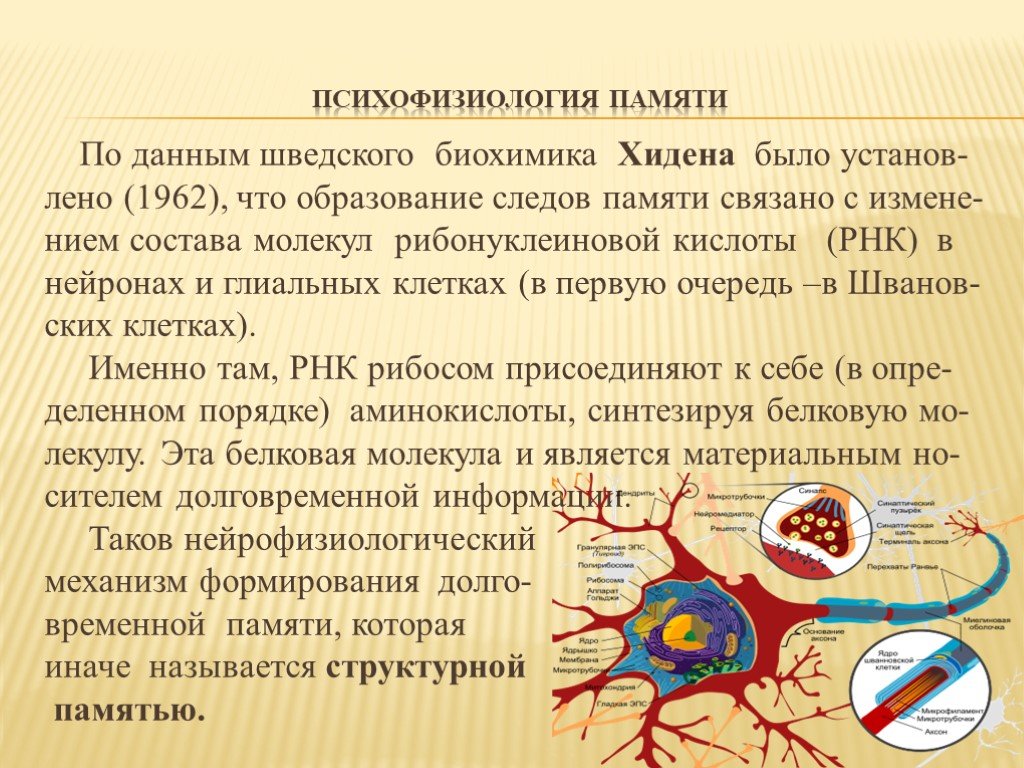
Теория памяти человека, зачатки ИИ / Хабр
Теория памяти человека, зачатки ИИ
Наверняка всем Вам очень хорошо известны такие моменты, когда нужно что-то вспомнить, но извлечь информацию из мозга становится большим пазлом.
Почему же такое происходит. Для начала немного теории работы нейрона, можно почитать тут или тут
Предположим, а может так оно и есть, все нейроны объединены в одни очень большой граф со сложной структурой. Данная структура сложна и не может работать хаотично, т.е. передаваемые импульсы передаются строго в определённом порядке, поэтому тут есть 2 варианта:
- Ребра графа имеют только положительные веса
- Ребра графа могут иметь, как положительные так и отрицательные веса
Что же касается первого случая, когда все ребра имеют положительные веса, т. е. головной мозг человека не поврежден. Тогда почему же человек не может вспомнить моменты из своего детства? Ответ прост: “Любое тело стремится к покою”, так же и наша с вами нейронная сеть старается оптимизировать свою работу. (Владельцам навигаторов должно быть знакомо, что прокладка маршрута, как раз таки строится на принципах работы графа, нахождения кратчайшего пути и т.д.). Мозг человека более изощренная система и его оптимизация заключается в разрыве связей с малыми весами, и построении новых связей с более высокими. (рис. 2). Таким образом объяснятся многочисленные разрывы и новые соединения нейронов. Чем больше узел имеет связей, тем легче вспомнить необходимую информацию.
е. головной мозг человека не поврежден. Тогда почему же человек не может вспомнить моменты из своего детства? Ответ прост: “Любое тело стремится к покою”, так же и наша с вами нейронная сеть старается оптимизировать свою работу. (Владельцам навигаторов должно быть знакомо, что прокладка маршрута, как раз таки строится на принципах работы графа, нахождения кратчайшего пути и т.д.). Мозг человека более изощренная система и его оптимизация заключается в разрыве связей с малыми весами, и построении новых связей с более высокими. (рис. 2). Таким образом объяснятся многочисленные разрывы и новые соединения нейронов. Чем больше узел имеет связей, тем легче вспомнить необходимую информацию.
Пример на практике
Вы сходили в кино, на обычный фильм, первое время вы помните практически все моменты, особенно те которые Вас как то эмоционально потрясли. Спустя месяц, год, у кого на сколько загружена деятельность мозга, если Вас спросить про этот фильм (с учетом того, что вы его не смотрели всё это время) Вы сможете рассказать краткий сюжет фильма, и те моменты которые Вас эмоционально потрясли. Это и есть оптимизация нейронов.
Это и есть оптимизация нейронов.Необходимо отметить, что каждый узел имеет свою пропускную способность, т.е. если идет нехватка связей, информацию можно и не вспомнить до необходимого перестроения графа. При оптимизации, передающийся импульс имеет более сильный сигнал, а сам узел свою пропускную способность не меняет, либо не значительно, именно поэтому очень важные вещи мы помним, как говорят на уровне подсознания.
В процессе воспоминания, Вы можете почувствовать недостаток информации, и продолжить вспоминать, тогда Ваш мозг ищет более длинный путь в графе (с более меньшим весом), и проходит по его граням в узлы которые в обычной жизни “не задействуются”, т.к. недостаточно накопленных импульсов для дальнейшей передачи сигнала.
Именно поэтому самый лучший способ воспоминания это ассоциации. Чем больше ребер в графе подходит к вершине, тем легче вспомнить информацию.
Оптимизация или частичная потеря воспоминаний
СОН – переключение мозга с активного запоминания в активную оптимизацию, во сне мы видим, то что уже когда-то видели либо синтез этих вещей. Синтез в данном случае это разрыв связей и возможно построение новой связи с другой вершиной.
Синтез в данном случае это разрыв связей и возможно построение новой связи с другой вершиной.Можно попробовать дать объяснение почему перед сном лучше запоминается. Во первых информация свежая, и достаточно полная (не оптимизированная), и наш мозг считает ее важной, придает вес. Во время ночной оптимизации информация полученная незадолго до сна, не успевает перемешаться с другой информацией и потеряется в “бесконечном” графе, а значит связи ее остаются практически неизменными. Конечно это только теория, у каждого человека мозг работает по своему, у каждого свои весы “полезной” и “бесполезной” информации.
Правила ИИ построенной на данной теории
- Вся накопленная информация за промежуток времени должна быть оптимизирована, т.е. установить причинно-следственную связь напрямую из начальной точки в конечные, пропуская промежуточные (разрываем сеть, и создаем новые связи с большим весом).
- После оптимизации все данные с незначительным весом должны быть уничтожены, а все веса понижены на минимальный вес из всех уничтоженных вершин.

- Новая информация должна иметь вес больше минимального существующего для правильной оценки важности информации.
- Всяка уничтоженная информация должна строить новый граф, и оптимизироваться. Причина тому, что постоянная оптимизация основного графа приведет к тому, не останется деталей, которые играют важную роль в жизни человека(эмоции).
Проблемы
В данном случае проблема состоит в оптимизации, т.е. выборке той информации которая будет жить, наиболее весомой для жизни человека и всей планеты, и той которая должна уйти на второй, третий, четвертый и т.д. план.Нейронная теория подсчета воспоминаний
Сохранить цитату в файл
Формат: Резюме (текст)PubMedPMIDAbstract (текст)CSV
Добавить в коллекции
- Создать новую коллекцию
- Добавить в существующую коллекцию
Имя должно содержать менее 100 символов
Выберите коллекцию: Не удалось загрузить вашу коллекцию из-за ошибки
Повторите попытку
Добавить в мою библиографию
- Моя библиография
Не удалось загрузить делегатов из-за ошибки
Повторите попытку
Ваш сохраненный поиск
Название сохраненного поиска:
Условия поиска:
Тестовые условия поиска
Электронная почта: (изменить)
Который день? Первое воскресеньеПервый понедельникПервый вторникПервая средаПервый четвергПервая пятницаПервая субботаПервый деньПервый рабочий день
Который день? ВоскресеньеПонедельникВторникСредаЧетвергПятницаСуббота
Формат отчета: РезюмеРезюме (текст)АбстрактАбстракт (текст)PubMed
Отправить максимум:
1 шт. 5 шт. 10 шт. 20 шт. 50 шт. 100 шт. 200 шт.
5 шт. 10 шт. 20 шт. 50 шт. 100 шт. 200 шт.
Отправить, даже если нет новых результатов
Необязательный текст в электронном письме:
Создайте файл для внешнего программного обеспечения для управления цитированием
. 2022 10 октября; 13 (1): 5961. doi: 10.1038/s41467-022-33577-2.Санджой Дасгупта 1 , Дайсуке Хаттори 2 , Сакет Навлаха 3
Принадлежности
- 1 Факультет компьютерных наук и инженерии, Калифорнийский университет в Сан-Диего, Ла-Хойя, Калифорния, 92037, США.
- 2 Отделение физиологии, Юго-Западный медицинский центр штата Юта, Даллас, Техас, 75390, США.
 [email protected].
[email protected]. - 3 Центр количественной биологии Симонса, лаборатория Колд Спринг Харбор, Колд Спринг Харбор, Нью-Йорк, 11724, США. [email protected].
- PMID: 36217003
- PMCID: PMC9551066
- DOI: 10.1038/с41467-022-33577-2
Санджой Дасгупта и др. Нац коммун. .
Бесплатная статья ЧВК . 2022 10 октября; 13 (1): 5961. doi: 10.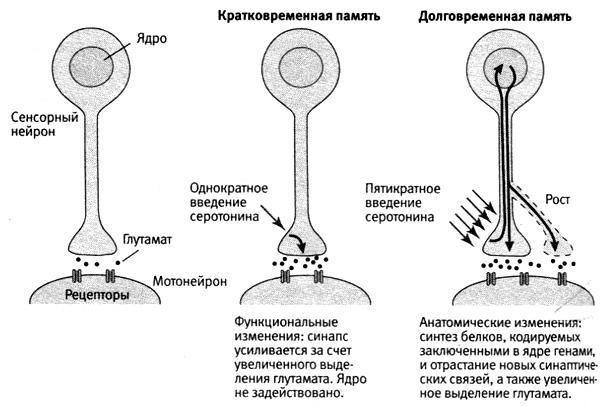 1038/s41467-022-33577-2.
1038/s41467-022-33577-2.Авторы
Санджой Дасгупта 1 , Дайсуке Хаттори 2 , Сакет Навлаха 3
Принадлежности
- 1 Факультет компьютерных наук и инженерии, Калифорнийский университет в Сан-Диего, Ла-Хойя, Калифорния, 92037, США.
- 2 Отделение физиологии, Юго-западный медицинский центр штата Юта, Даллас, Техас, 75390, США. [email protected].
- 3 Центр количественной биологии Симонса, лаборатория Колд Спринг Харбор, Колд Спринг Харбор, Нью-Йорк, 11724, США. navlakha@cshl.
 edu.
edu.
- PMID: 36217003
- PMCID: PMC9551066
- DOI: 10.1038/с41467-022-33577-2
Абстрактный
Отслеживание того, сколько раз были испытаны различные стимулы, является критическим вычислением поведения. Здесь мы предлагаем теоретическую двухуровневую нейронную цепь, в которой хранятся подсчеты частот появления стимулов. Эта схема реализует структуру данных, называемую схемой подсчета, которая обычно используется в компьютерных науках для поддержания частоты элементов в потоковых данных. Наша первая модель реализует эскиз счета с использованием синапсов Хебба и выводит частоты, специфичные для стимула.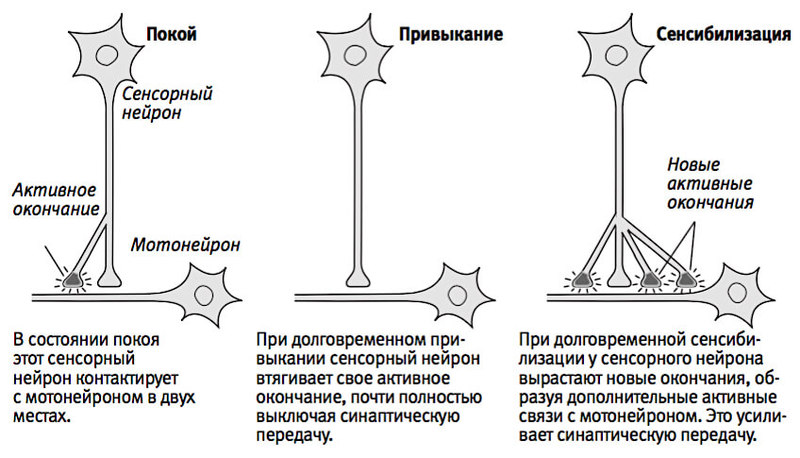 Наша вторая модель использует антихеббовскую пластичность и отслеживает частоты только в пределах четырех категорий счета («1-2-3-много»), что приводит к компромиссу количества категорий, которые необходимо различать, с потенциальной этологической ценностью этих категорий. Мы показываем, как обе модели могут надежно отслеживать частоты появления стимулов, тем самым расширяя традиционную ось памяти новизна-знакомство с бинарной на дискретную с более чем двумя возможными значениями. Наконец, мы показываем, что реализация схемы подсчета «1-2-3-много» существует в грибовидном теле насекомого.
Наша вторая модель использует антихеббовскую пластичность и отслеживает частоты только в пределах четырех категорий счета («1-2-3-много»), что приводит к компромиссу количества категорий, которые необходимо различать, с потенциальной этологической ценностью этих категорий. Мы показываем, как обе модели могут надежно отслеживать частоты появления стимулов, тем самым расширяя традиционную ось памяти новизна-знакомство с бинарной на дискретную с более чем двумя возможными значениями. Наконец, мы показываем, что реализация схемы подсчета «1-2-3-много» существует в грибовидном теле насекомого.
© 2022. Автор(ы).
Заявление о конфликте интересов
Авторы заявляют об отсутствии конкурирующих интересов.
Цифры
Рис. 1. Эскиз графа и соответствующий…
1. Эскиз графа и соответствующий…
Рис. 1. Эскиз счетчика и соответствующая реализация нейронной схемы.
A Данные эскиза счета…
Рис. 1. Эскиз счетчика и соответствующая реализация нейронной схемы. A Структура данных эскиза счета представляет собой двумерную матрицу C счетчиков с тыс. строк и против столбцов. Существует одна хеш-функция х на строку, каждая из которых определяет, какой столбец в строке изменяется при наблюдении элемента размером х (пунктирные стрелки). B Нейронная реализация схемы подсчета использует одномерный массив w из k × v синапсов. Когда объект наблюдается, синапсы k пресинаптических нейронов, которые активны для объекта (выделено оранжевым цветом), изменяются. Нейроны, активированные для элемента, определяются с помощью хэш-функции, которая присваивает элементу разреженное многомерное представление ( z ). В этом примере каждый элемент размером 90 177 x 90 178 представляет собой 90 177 d 90 178 -мерный вектор. C Чтобы вставить элемент из последовательности в схему подсчета (вверху), k счетчиков увеличиваются. Например, после первого наблюдения ( x 1 ) в первой строке счетчик в 1-м столбце (т. е. C [1, 1]) увеличивается на 1, поскольку 7 x 1 ) = 1. ) = 2. В третьем ряду C [3, 5] увеличивается на 1, так как h 3 ( x 1 ) = 5. k активированных пресинаптических нейронов для x 1 (выделено оранжевым цветом) увеличиваются. В антихеббовской модели все синаптические веса инициализируются равными 1, а синапсы, активные для элемента, уменьшаются при каждом наблюдении. D Для вывода оценки частоты элемента x 1 , скетч подсчета вычисляет: (1/k)∑i=1kC[i,hi(x1)] — т.е. среднее предсказанных подсчетов по строкам. Это приводит к правильным оценкам для x 1 и x 2 и почти правильной оценке для x 3 .
В этом примере каждый элемент размером 90 177 x 90 178 представляет собой 90 177 d 90 178 -мерный вектор. C Чтобы вставить элемент из последовательности в схему подсчета (вверху), k счетчиков увеличиваются. Например, после первого наблюдения ( x 1 ) в первой строке счетчик в 1-м столбце (т. е. C [1, 1]) увеличивается на 1, поскольку 7 x 1 ) = 1. ) = 2. В третьем ряду C [3, 5] увеличивается на 1, так как h 3 ( x 1 ) = 5. k активированных пресинаптических нейронов для x 1 (выделено оранжевым цветом) увеличиваются. В антихеббовской модели все синаптические веса инициализируются равными 1, а синапсы, активные для элемента, уменьшаются при каждом наблюдении. D Для вывода оценки частоты элемента x 1 , скетч подсчета вычисляет: (1/k)∑i=1kC[i,hi(x1)] — т.е. среднее предсказанных подсчетов по строкам. Это приводит к правильным оценкам для x 1 и x 2 и почти правильной оценке для x 3 . Точно так же скетч нейронного подсчета выводит: (1/k)∑i,zi>0wizi — т. е. среднее значение весов активированных нейронов для элемента.
Точно так же скетч нейронного подсчета выводит: (1/k)∑i,zi>0wizi — т. е. среднее значение весов активированных нейронов для элемента.
Рис. 2. Производительность нейронного счета…
Рис. 2. Выполнение скетча нейронного подсчета (модель Хебба).
В каждой панели x…
Рис. 2. Выполнение скетча нейронного подсчета (модель Хебба). На каждой панели ось x показывает подсчет истинности, а ось y показывает предсказанный подсчет, выдаваемый нейроном декодера. Точки показывают средние ответы декодера для элементов с одинаковым числом достоверных данных, а планки погрешностей указывают на стандартное отклонение. Идеальная производительность будет лежать на пунктирной г = x строка. Коэффициент корреляции Пирсона ( r ) количественно определяет точность производительности (чем больше, тем лучше). После удаления связанных элементов N = 1000 для синтетических материалов; N = 62 для запахов; и N = 180 для MNIST. Каждая панель показывает набор данных (столбцы) с добавлением шума к элементам (строкам) или без него. A , B Синтетика без шума и с шумом. C , D Запахи без шума и с шумом. Е , F MNIST без шума и с шумом.
После удаления связанных элементов N = 1000 для синтетических материалов; N = 62 для запахов; и N = 180 для MNIST. Каждая панель показывает набор данных (столбцы) с добавлением шума к элементам (строкам) или без него. A , B Синтетика без шума и с шумом. C , D Запахи без шума и с шумом. Е , F MNIST без шума и с шумом.
Рис. 3. Выполнение скетча 1-2-3-many…
Рис. 3. Выполнение скетча 1-2-3-много (антихеббовская модель).
В каждой панели ось x …
Рис. 3. Выполнение скетча 1-2-3-many (антихеббовская модель). На каждой панели ось x показывает четыре категории подсчета истинности, а ось y показывает выходные данные декодера из эскиза 1-2-3-множество. Например, категория счета «1» включает в себя все элементы, наблюдаемые впервые (т. е. элементы, отсутствующие во входной последовательности), а выходные данные декодера показывают величину отклика нейрона декодера для всех таких элементов. Блочные диаграммы показывают медиану, а также первый и третий квартили, при этом усы простираются от прямоугольника в 1,5 раза больше межквартильного диапазона. Вверху показаны p -значения (критерий суммы рангов Уилкоксона, двусторонний), сравнивающие различия в величинах ответов декодера между элементами в последовательных категориях подсчета. Например, на панели А разница между ответами декодера на элементы в счетной категории «1» и «2» была статистически значимой, с 90 177 p 90 178 = 2,1e-120. Все p < 0,01 окрашены в красный цвет. После удаления связанных элементов N = 1000 для синтетических материалов; N = 62 для запахов; и N = 180 для MNIST. Каждая панель показывает набор данных (столбцы) с добавлением шума к элементам (строкам) или без него. A , B Синтетика без шума и с шумом.
е. элементы, отсутствующие во входной последовательности), а выходные данные декодера показывают величину отклика нейрона декодера для всех таких элементов. Блочные диаграммы показывают медиану, а также первый и третий квартили, при этом усы простираются от прямоугольника в 1,5 раза больше межквартильного диапазона. Вверху показаны p -значения (критерий суммы рангов Уилкоксона, двусторонний), сравнивающие различия в величинах ответов декодера между элементами в последовательных категориях подсчета. Например, на панели А разница между ответами декодера на элементы в счетной категории «1» и «2» была статистически значимой, с 90 177 p 90 178 = 2,1e-120. Все p < 0,01 окрашены в красный цвет. После удаления связанных элементов N = 1000 для синтетических материалов; N = 62 для запахов; и N = 180 для MNIST. Каждая панель показывает набор данных (столбцы) с добавлением шума к элементам (строкам) или без него. A , B Синтетика без шума и с шумом.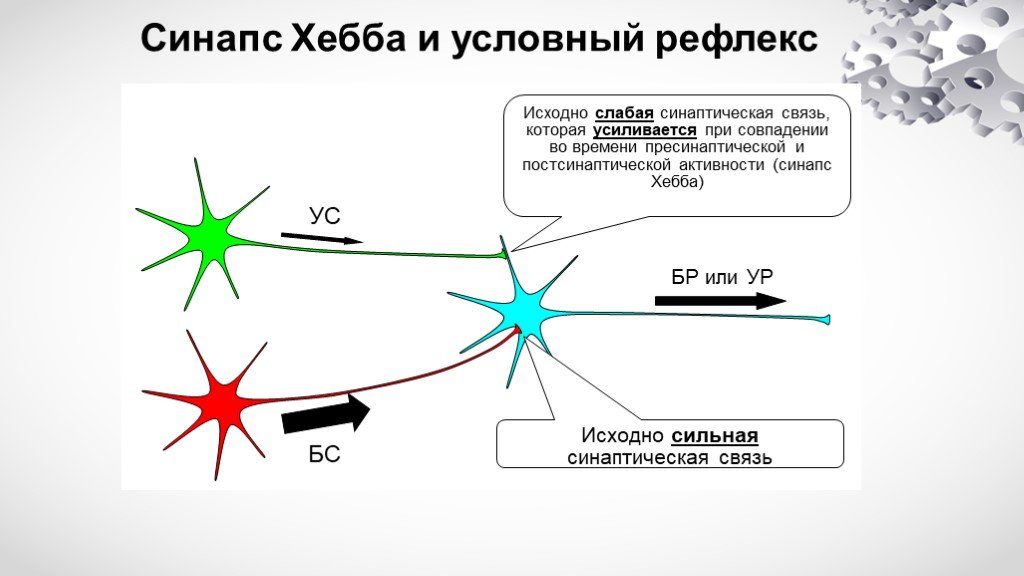 C , D Запахи без шума и с шумом. E , F MNIST без шума и с шумом.
C , D Запахи без шума и с шумом. E , F MNIST без шума и с шумом.
Рис. 4. Экспериментальное свидетельство «1-2-3-много»…
Рис. 4. Экспериментальное свидетельство эскиза «1-2-3-много» из грибовидного тела насекомого.
A Схема…
Рис. 4. Экспериментальное свидетельство эскиза «1-2-3-много» с тела гриба насекомого. A Схема обонятельной системы плодовой мушки. Запахи изначально представлены скоростью возбуждения 50 типов нейронов рецепторов запахов, которые посылают аксоны к 50 типам проекционных нейронов (PN). Затем PN отправляют информацию о запахе в 2000 клеток Кеньона (KC), каждая из которых обеспечивает прямое возбуждение большого тормозного нейрона (называемого APL), который разрежает представление KC посредством торможения по обратной связи. B Синапсы между активированными KC и счетным нейроном (MBON-α’3) модифицируются (ослабевают) при восприятии запаха. C Динамика реакции MBON-α′3 ( и по оси) на 10 последовательных предъявлений запаха MCH ( x по оси). Данные показывают ответы N = 72 клеток (светло-голубой) на 59 мухах. Синие точки (темно-синие) показывают медианные значения отклика, а планки ошибок показывают 99% доверительные интервалы, определенные с помощью 20 000 бутстрапов. Для каждой клетки ответы нормированы на величину первого представления. Красная кривая показывает данные, соответствующие экспоненциальной функции ( y = a e b x ), с константой подавления 0,44. Исходные данные предоставляются в виде файла исходных данных. D Тепловая карта p -значений (критерий суммы рангов Уилкоксона, двусторонний), сравнивающая различия в величинах ответов для всех пар категорий подсчета. Например, ответы MBON-α’3 значительно отличаются при сравнении запаха, увиденного в первый раз, и во второй раз ( p = 1,9e-23), но ответы существенно не отличаются по сравнению с 4-м и 5-м опытом (9).
B Синапсы между активированными KC и счетным нейроном (MBON-α’3) модифицируются (ослабевают) при восприятии запаха. C Динамика реакции MBON-α′3 ( и по оси) на 10 последовательных предъявлений запаха MCH ( x по оси). Данные показывают ответы N = 72 клеток (светло-голубой) на 59 мухах. Синие точки (темно-синие) показывают медианные значения отклика, а планки ошибок показывают 99% доверительные интервалы, определенные с помощью 20 000 бутстрапов. Для каждой клетки ответы нормированы на величину первого представления. Красная кривая показывает данные, соответствующие экспоненциальной функции ( y = a e b x ), с константой подавления 0,44. Исходные данные предоставляются в виде файла исходных данных. D Тепловая карта p -значений (критерий суммы рангов Уилкоксона, двусторонний), сравнивающая различия в величинах ответов для всех пар категорий подсчета. Например, ответы MBON-α’3 значительно отличаются при сравнении запаха, увиденного в первый раз, и во второй раз ( p = 1,9e-23), но ответы существенно не отличаются по сравнению с 4-м и 5-м опытом (9). 0177 p = 5.3e-02). Красные блоки обозначают p < 0,01, а серые блоки указывают p ≥ 0,01. E Распределение гистограмм ответов MBON-α′3 для каждой категории счета, с кривыми оценки плотности ядра, нанесенными сверху. Категории 1, 2, 3 и 4 показаны синим, оранжевым, красным и зеленым цветом соответственно; категории 5–10 показаны черным цветом. Категории 4–10 (многие) сильно перекрываются и отличаются от категорий 1, 2 и 3. F Тепловая карта области под значениями ROC (auROC) для различения каждой пары категорий количества с использованием линейного порога, применяемого к ответу MBON-α′3. Например, auROC для различения 1-против-2 составлял 0,9.82. Отличительность является самой высокой для категорий 1, 2 и 3 (первые три столбца), а затем снижается для последующих категорий.
0177 p = 5.3e-02). Красные блоки обозначают p < 0,01, а серые блоки указывают p ≥ 0,01. E Распределение гистограмм ответов MBON-α′3 для каждой категории счета, с кривыми оценки плотности ядра, нанесенными сверху. Категории 1, 2, 3 и 4 показаны синим, оранжевым, красным и зеленым цветом соответственно; категории 5–10 показаны черным цветом. Категории 4–10 (многие) сильно перекрываются и отличаются от категорий 1, 2 и 3. F Тепловая карта области под значениями ROC (auROC) для различения каждой пары категорий количества с использованием линейного порога, применяемого к ответу MBON-α′3. Например, auROC для различения 1-против-2 составлял 0,9.82. Отличительность является самой высокой для категорий 1, 2 и 3 (первые три столбца), а затем снижается для последующих категорий.
Рис. 5. Предполагаемые механизмы считывания…
Рис. 5. Гипотетические механизмы считывания счетного нейрона.
5. Гипотетические механизмы считывания счетного нейрона.
A Эта модель переводит непрерывную…
Рис. 5. Гипотетические механизмы считывания счетного нейрона. A Эта модель переводит непрерывную частоту срабатывания счетчика 1-2-3-множество нейронов в дискретное представление. Поскольку в этой модели используется подавление повторения, более высокая скорость срабатывания соответствует менее знакомым запахам. Счетчик синапсирует с четырьмя нисходящими нейронами, каждый из которых имеет последовательно уменьшающиеся пороги срабатывания ( θ 1 > θ 2 > θ 3 > θ 4 ). Нейроны с более высокими порогами подавляют нейроны с меньшими порогами. В результате каждая категория счетчика становится «горячей» закодированной, что упрощает изменение поведения. B Эта модель иерархически связывает вместе счетные нейроны для увеличения масштабируемости и разрешения между категориями счета. В грибовидном теле насекомого запахи активируют дофаминовые нейроны (ДАН), которые модулируют синаптические веса счетных нейронов. В этой модели есть два счетных нейрона с двумя связанными DAN. Оба счетчика получают входные данные от кодирующих нейронов, а Counter1 подавляет Counter2 и DAN2. Counter1 кодирует категории 1 и 2 с высокой (1,0) и средней (0,5) чувствительностью соответственно. Когда активность Счетчика 1 уменьшается после второго воздействия стимула, торможение Счетчика 2 снимается, позволяя Счетчику 2 кодировать две последующие категории с высокой и средней реакцией.
В грибовидном теле насекомого запахи активируют дофаминовые нейроны (ДАН), которые модулируют синаптические веса счетных нейронов. В этой модели есть два счетных нейрона с двумя связанными DAN. Оба счетчика получают входные данные от кодирующих нейронов, а Counter1 подавляет Counter2 и DAN2. Counter1 кодирует категории 1 и 2 с высокой (1,0) и средней (0,5) чувствительностью соответственно. Когда активность Счетчика 1 уменьшается после второго воздействия стимула, торможение Счетчика 2 снимается, позволяя Счетчику 2 кодировать две последующие категории с высокой и средней реакцией.
См. это изображение и информацию об авторских правах в PMC
Похожие статьи
- Эффективные ассоциативные вычисления с дискретными синапсами.
Кноблаух А. Кноблаух А. Нейронные вычисления. 2016 Январь; 28 (1): 118-86. дои: 10.1162/NECO_a_00795. Epub 2015 24 ноября.
 Нейронные вычисления. 2016.
PMID: 26599711
Нейронные вычисления. 2016.
PMID: 26599711 - Модель нейронной сети для ознакомления и контекстного обучения во время полетов медоносных пчел за кормом.
Мюллер Дж., Наврот М., Менцель Р., Ландграф Т. Мюллер Дж. и др. Биол Киберн. 2018 апрель; 112 (1-2): 113-126. doi: 10.1007/s00422-017-0732-z. Epub 2017 15 сентября. Биол Киберн. 2018. PMID: 28917001
- Сравнение вычислительных моделей распознавания знакомств в околоносовой области коры.
Богач Р., Браун М.В. Богач Р. и соавт. Гиппокамп. 2003;13(4):494-524. doi: 10.1002/hipo.10093. Гиппокамп. 2003. PMID: 12836918 Обзор.
- Стабильная память и вычисления в случайно перемонтируемых нейронных сетях.

Акер Д., Паради С., Миллер П. Акер Д. и соавт. J Нейрофизиол. 2019 1 июля; 122 (1): 66-80. doi: 10.1152/jn.00534.2018. Эпаб 201910 апр. J Нейрофизиол. 2019. PMID: 30969897 Бесплатная статья ЧВК.
- Хеббовская пластичность требует компенсационных процессов в нескольких временных масштабах.
Зенке Ф., Герстнер В. Зенке Ф. и др. Philos Trans R Soc Lond B Biol Sci. 2017 5 марта; 372 (1715): 20160259. doi: 10.1098/rstb.2016.0259. Philos Trans R Soc Lond B Biol Sci. 2017. PMID: 28093557 Бесплатная статья ЧВК. Обзор.
Посмотреть все похожие статьи
Рекомендации
- Jaegle A, Mehrpour V, Rust N. Визуальная новизна, любопытство и внутреннее вознаграждение в машинном обучении и мозге.
 Курс. мнение Нейробиол. 2019; 58: 167–174. doi: 10.1016/j.conb.2019.08.004.
—
DOI
—
пабмед
Курс. мнение Нейробиол. 2019; 58: 167–174. doi: 10.1016/j.conb.2019.08.004.
—
DOI
—
пабмед
- Jaegle A, Mehrpour V, Rust N. Визуальная новизна, любопытство и внутреннее вознаграждение в машинном обучении и мозге.
- Браун М.В., Эглтон Дж.П. Память узнавания: каковы роли околоносовой коры и гиппокампа? Нац. Преподобный Нейроски. 2001; 2:51–61. дои: 10.1038/35049064. — DOI — пабмед
- Десимоне Р.
 Нейронные механизмы зрительной памяти и их роль во внимании. проц. Натл акад. науч. США. 1996; 93:13494–13499. doi: 10.1073/pnas.93.24.13494.
—
DOI
—
ЧВК
—
пабмед
Нейронные механизмы зрительной памяти и их роль во внимании. проц. Натл акад. науч. США. 1996; 93:13494–13499. doi: 10.1073/pnas.93.24.13494.
—
DOI
—
ЧВК
—
пабмед
- Десимоне Р.
- Браун М.В., Сян Дж.З. Память узнавания: нейронные субстраты суждения о предыдущем происшествии. прог. Нейробиол. 1998; 55: 149–189. doi: 10.1016/S0301-0082(98)00002-1. — DOI — пабмед
- Сквайр Л.
 Р., Шмолк Х., Старк С.М. Нарушение слуховой памяти у пациентов с амнезией и медиальными поражениями височной доли. Учиться. Мем. 2001; 8: 252–256. doi: 10.1101/lm.42001.
—
DOI
—
ЧВК
—
пабмед
Р., Шмолк Х., Старк С.М. Нарушение слуховой памяти у пациентов с амнезией и медиальными поражениями височной доли. Учиться. Мем. 2001; 8: 252–256. doi: 10.1101/lm.42001.
—
DOI
—
ЧВК
—
пабмед
- Сквайр Л.
Типы публикаций
термины MeSH
Грантовая поддержка
- R01 DC017695/DC/NICDD NIH HHS/США
- UF1 NS111692/NS/NINDS NIH HHS/США
Укажите
Формат: ААД АПА МДА НЛМ
Отправить по номеруМатематическая теория нейронных сетей объясняет, как воспоминания объединяются в мозгу
- Скачать PDF Копировать
Отзыв от Lily Ramsey, LLM 20 июля 2023
То, насколько полезна память для будущих ситуаций, определяет, где она находится в мозгу, согласно новой теории, предложенной исследователями исследовательского кампуса Джанелия HHMI и сотрудниками UCL.
Теория предлагает новый способ понимания системной консолидации, процесса, при котором определенные воспоминания передаются из гиппокампа, где они изначально хранятся, в неокортекс, где они хранятся долгое время.
Согласно классическому взгляду на консолидацию систем, все воспоминания со временем перемещаются из гиппокампа в неокортекс. Но эта точка зрения не всегда верна; исследования показывают, что некоторые воспоминания постоянно находятся в гиппокампе и никогда не передаются в неокортекс.
В последние годы психологи предложили теории, объясняющие этот более сложный взгляд на консолидацию систем, но никто еще не вычислил математически, что определяет, остается ли память в гиппокампе или консолидируется в неокортексе.
Теперь исследователи Janelia выдвигают новый, количественный взгляд на консолидацию систем, чтобы помочь решить эту давнюю проблему, предлагая математическую теорию нейронных сетей, в которой воспоминания консолидируются в неокортексе только в том случае, если они улучшают обобщение.
Обобщения строятся на основе надежных и предсказуемых компонентов воспоминаний, что позволяет нам применять их в других ситуациях. Мы можем обобщить некоторые особенности воспоминаний, чтобы помочь нам понять мир, например, тот факт, что каньоны предсказывают присутствие воды.
Related Stories
- Новый сверхгибкий нейронный имплант регистрирует активность одиночных нейронов в глубоких отделах мозга
- Исследование изучает черепно-мозговую травму у детей как потенциальный фактор риска психотических расстройств или симптомов
- Ориентация на сигнал «не ешь меня» может улучшить иммунотерапию глиобластомы
Это отличается от эпизодических воспоминаний — подробных воспоминаний о прошлом, которые имеют уникальные особенности, такие как индивидуальное воспоминание о походе в определенный каньон и встрече с водоемом.
С этой точки зрения консолидация не копирует воспоминания из одной области мозга в другую, а скорее создает новую память, которая является обобщением предыдущих воспоминаний. Объем воспоминаний, а не их возраст, определяет, консолидируются они или остаются в гиппокампе.
Объем воспоминаний, а не их возраст, определяет, консолидируются они или остаются в гиппокампе.
Исследователи использовали нейронные сети, чтобы показать, как степень консолидации варьируется в зависимости от того, насколько обобщаема память. Они смогли воспроизвести предыдущие экспериментальные модели, которые нельзя было объяснить с классической точки зрения на консолидацию систем.
Следующий шаг — проверить теорию экспериментами, чтобы увидеть, может ли она предсказать, насколько память будет консолидирована. Другим важным направлением будет проверка моделей авторов того, как мозг может различать предсказуемые и непредсказуемые компоненты воспоминаний, чтобы регулировать консолидацию. Раскрытие того, как работает память, может помочь исследователям лучше понять неотъемлемую часть познания, что может принести пользу здоровью человека и искусственному интеллекту.
Источник:
Медицинский институт Говарда Хьюза
Ссылка на журнал:
Sun, W.
